기생 (생물학)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
기생은 다른 생물체(숙주)의 표면 또는 내부에 서식하며 숙주로부터 영양을 섭취하거나 숙주에게 해를 끼치는 생물학적 현상을 의미한다. 기생은 숙주의 표면에 기생하는 외부 기생과 숙주 내부에 기생하는 내부 기생으로 나뉘며, 내부 기생은 세포간 기생과 세포내 기생으로 구분된다. 기생충은 숙주와의 관계, 기생 위치, 생활사에 따라 다양하게 분류되며, 기생충은 숙주의 방어 기제를 극복하고 전파 전략을 통해 생존한다. 기생은 생태계에서 중요한 역할을 하며, 숙주와 기생충은 공진화를 통해 서로 영향을 주고받는다. 한국은 과거 기생충 질환 유병률이 높았으나, 국가적인 기생충 관리 사업을 통해 크게 감소했다.
기생은 공생의 한 종류로, 기생충과 숙주 사이의 긴밀하고 지속적인 장기적인 생물학적 상호 작용이다. 부생생물과 달리 기생충은 살아있는 숙주를 먹고 살지만, 일부 기생 곰팡이는 숙주를 죽인 후에도 계속 먹고 살 수 있다. 공생과 상리공생과 달리, 기생 관계는 숙주에 해를 끼치며, 숙주를 먹거나 장내 기생충의 경우처럼 숙주의 일부 음식을 소비한다. 기생충은 다른 종과 상호 작용하기 때문에 쉽게 질병 매개체 역할을 하여 기생충 질환을 유발할 수 있다.[6][7][8]
2. 기생의 정의 및 유형
기생에는 다음과 같은 6가지 주요 전략이 있다.[19][20]
1539년 영어에서 처음 사용된 ''parasite''라는 단어는 중세 프랑스어 parasitefro에서 유래했으며, 이는 라틴화된 형태 parasitusla에서 비롯되었다.[4] 관련된 용어인 ''parasitism''은 1611년부터 영어에 등장한다.[5]
기생은 다양한 기준에 따라 분류될 수 있다.
2. 1. 기생 위치에 따른 분류
진드기(mite)나 머릿니처럼 숙주의 표면에 붙어 사는 기생충들을 '''외부기생충'''(ectoparasites)이라고 한다.[12] 
반대로 장내 기생충을 포함한 모든 숙주 내부에 사는 것들을 '''내부기생충'''이라고 한다. 내부기생충 내에서도 '''세포간 기생'''(intercellular)과 '''세포내 기생'''(intracellular)이라는 두 가지 생존 방식이 있다. 세포간 기생을 하는 세균이나 바이러스들은 보균체(carrier or vector)라 불리는 제3의 개체를 필요로 하는 경우가 있다. 보균체는 기생충을 숙주에 옮기는 역할을 맡는다. 말라리아의 감염 경로를 예로 들 수 있는데, 인간이 말라리아 모기에 물리면 모기 내에 있던 말라리아 속 원생동물이 체내로 유입되어 말라리아에 감염된다.[12]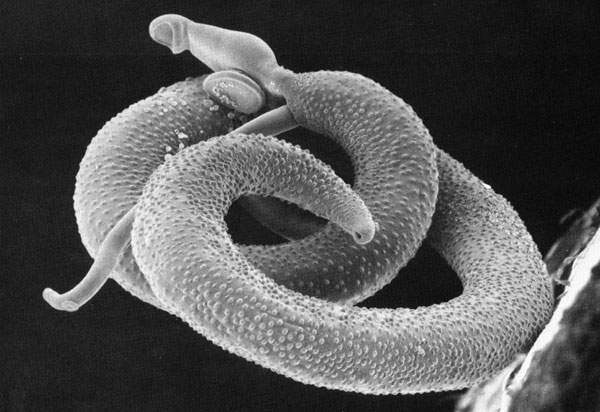
기생자가 숙주의 세포 내에 들어가 있는 경우, '''세포 내 기생'''이라고 한다. 말라리아 원충이 그 예이다.
어디까지가 외부이고 어디부터가 내부인지에 대해서는 약간의 논의가 있다. 예를 들어 벼룩은 명백히 외부 기생이다. 그러나 모래벼룩 암컷은 한 곳을 물면 거기에서 움직이지 않고, 주변부의 피부가 융기하여 묻혀 버린다. 이것은 내부 기생이라고 하는 경우도 있다. 기생성 파리목 유충에는 상처 부위에 잠입하여 꼬리를 밖으로 내는 것이 있다. 소화관 안에 생식하는 기생충은 내부 기생이라고 하는 것이 보통이지만, 비강 내의 것은 어떠한지, 연체동물의 외투막 내부는 어느 쪽인지 등 생각하면 어렵다. 엄밀히 말하면, 내부 기생은 체강 내에 생식하는 것과 세포 내 기생만을 포함해야 한다는 생각도 있다.
2. 2. 숙주와의 관계에 따른 분류
기생은 기본적으로 숙주와의 관계 그리고 그들의 생활 주기에 근거해 분류된다.
: 진드기(mite), 머릿니처럼 숙주의 표면에 붙어 사는 기생충들을 '''외부기생충'''(ectoparasites)이라고 한다.[12]
: 장내 기생충을 포함한 모든 숙주 내부에 사는 것들을 '''내부기생충'''이라고 한다. 내부기생충 내에서도 '''세포간 기생'''(intercellular)과 '''세포내 기생'''(intracellular)이라는 두 가지 생존 방식이 있다. 세포간 기생을 하는 세균이나 바이러스들은 보균체(carrier or vector)라 불리는 제3의 개체를 필요로 하는 경우가 있다. 보균체는 기생충을 숙주에 옮기는 역할을 맡는다. 말라리아의 감염 경로가 이러한 예인데, 인간이 말라리아 모기에 물리면 모기 내에 있던 말라리아 속 원생동물이 체내로 유입되어 말라리아에 감염된다.
: '''착생기생충'''(epiparasite)은 다른 기생충에 의지해 살아간다. 때로는 기생체내기생(hyperparasitism)이라고 불린다. 개에 기생해 사는 벼룩의 소화간에 기생하는 원생동물이 이에 해당한다.
: '''사회 기생충'''(social parasites)은 개미나 흰개미처럼 사회적 생활을 하는 개체들로부터 이득을 얻는다. '''도적 기생충'''(kleptoparasitism)들은 숙주가 구해온 먹이를 갈취한다. 조류에서 볼 수 있는 또 다른 사회 기생의 예로, 뻐꾸기나 찌르레기들은 둥지를 짓지 않고 다른 종의 둥지에 알을 놓고 도망가 버린다. 둥지의 주인은 숙주가 되어 버려진 알을 마치 자신의 알처럼 품어서 부화시킨다. 숙주가 보모의 역할을 하는 것과 같다. 만약 숙주가 뻐꾸기의 알을 버리면, 어떤 뻐꾸기들은 둥지로 가서 숙주를 괴롭혀 굴복시킨다. 뻐꾸기들은 가끔 들키지 않기 위해 숙주의 알 몇 개를 제거하기도 하고, 태어난 뻐꾸기들이 숙주의 알들을 떨어뜨리거나 새끼들을 괴롭히기도 한다. 반면에 찌르레기는 숙주에게 해를 가하지 않는다.
: '''포식 기생충'''(parasitoids)의 애벌레는 다른 개체의 체내나 체외에서 성장하며, 성장이 끝나는 순간 숙주는 죽음을 맞이한다. 그런 의미에서 포식 기생은 여타의 기생과는 구별되며, 포식관계와 비슷하다고 볼 수 있다.
분류학자들은 숙주와의 상호 작용과 복잡할 수 있는 생물학적 생활사를 기반으로 다양한 방식으로 기생충을 분류한다. 절대 기생충은 생활사를 완료하기 위해 숙주에 전적으로 의존하는 반면, 조건적 기생충은 그렇지 않다.[10][11]
2. 3. 생활사에 따른 분류
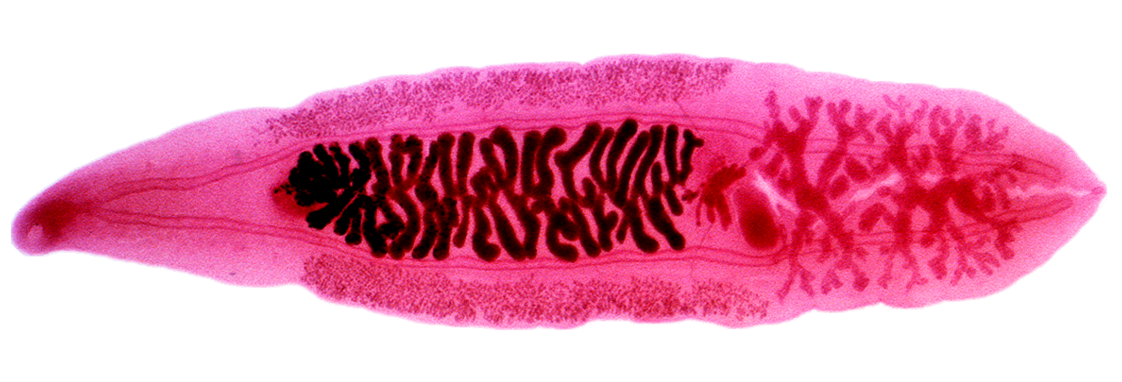
영양적으로 전파되는 기생충은 숙주에게 먹힘으로써 전파된다. 여기에는 흡충류(주혈흡충류 제외), 촌충, 극두동물, 설충, 많은 선충류, 그리고 ''톡소포자충''과 같은 많은 원생동물이 포함된다.[20] 이들은 두 종 이상의 숙주를 포함하는 복잡한 생물학적 생활사를 가지고 있다. 유충 단계에서 중간 숙주를 감염시키고 종종 중간 숙주 내에서 낭포를 형성한다. 중간 숙주 동물(최종 숙주)이 포식자에게 먹히면, 기생충은 소화 과정을 거쳐 살아남아 성충으로 성숙하며, 일부는 장내 기생충으로 산다. 많은 영양적으로 전파되는 기생충들은 중간 숙주의 행동을 수정하여 포식자에게 잡아먹힐 가능성을 높인다. 직접 전파되는 기생충과 마찬가지로, 영양적으로 전파되는 기생충의 숙주 개체 간 분포는 집합적이다.[20] 여러 기생충에 의한 중복 감염이 흔하다.[24] 예외적으로 기생충의 전체 생물학적 생활사가 단일 일차 숙주 내에서 발생하는 자가 감염은 ''분선충''과 같은 선충류에서 때때로 발생할 수 있다.[25]
2. 4. 기타 분류
사회 기생은 개미, 흰개미, 호박벌과 같이 사회적 생활을 하는 생물들 사이에서 나타나는 기생 형태이다. 예를 들어, 큰 파란나비(''Phengaris arion'')의 유충은 개미 모방을 통해 특정 개미에게 기생한다.[37] 호박벌( ''Bombus bohemicus'')은 다른 벌의 벌집을 침입하여 숙주 일벌에게 새끼를 키우게 하고 번식을 장악한다.[38] 또한, ''Melipona scutellaris''라는 진사회성 벌은 여왕이 없는 다른 군집에 침입하여 살해자 일벌을 탈출하는 방식으로 기생한다.[38] 이러한 사회적 기생의 극단적인 예로, 다른 ''Tetramorium'' 개미의 등에만 사는 절대 기생충인 개미 ''Tetramorium inquilinum''가 있다.[39]
사회 기생의 진화 메커니즘은 1909년 카를로 에머리에 의해 처음 제안되었으며,[40] 현재는 "에머리 규칙"으로 알려져 있다. 이 규칙은 사회적 기생충이 숙주와 밀접하게 관련되어 있으며, 종종 같은 속에 속한다고 명시한다.[41][42][43]
종내 사회적 기생의 경우, 일부 개체가 관련 없는 암컷에게서 젖을 섭취하는 기생적 간호가 발생하기도 한다. 예를 들어, 쐐기머리 카푸친에서는 더 높은 지위의 암컷이 낮은 지위의 암컷에게서 젖을 섭취하지만, 이에 대한 보답은 하지 않는다.[44]
포식 기생은 기생충의 애벌레가 다른 개체의 체내나 체외에서 성장하며, 성장이 끝날 때 숙주를 죽음에 이르게 하는 기생 형태이다. 이는 포식 관계와 유사하다고 볼 수 있다. 예를 들어 기생벌인 고치벌은 배추흰나비의 유충에 기생하는데, 고치벌 애벌레는 숙주인 배추흰나비의 몸속에서 기생하다가 뚫고 나와 고치를 만든다.
3. 숙주의 방어 기제

숙주는 기생충에 대항하기 위해 다양한 방어 기제를 진화시켜 왔다. 여기에는 척추동물의 피부와 같은 물리적 장벽,[106] 포유류의 면역 체계,[107] 곤충이 기생충을 적극적으로 제거하는 행위,[108] 식물의 방어 화학 물질 등이 포함된다.[109]
W. D. 해밀턴은 유성 생식이 유전자 재조합을 통해 다양한 유전자 조합을 생성하여 여러 기생충을 물리치는 데 도움이 되도록 진화했을 수 있다고 제안했다. 해밀턴은 수학적 모델링을 통해 유성 생식이 다양한 상황에서 진화적으로 안정적이며, 이론의 예측이 유성 생식의 실제 생태와 일치한다는 것을 보여주었다.[103][104] 그러나 면역 능력과 수컷 척추동물 숙주의 2차 성징 간에는 상충 관계가 있을 수 있다. 예를 들어 공작의 깃털이나 사자의 갈기는 수컷 호르몬인 테스토스테론이 2차 성징의 성장을 촉진하여 성 선택에서 유리하게 작용하지만, 면역 방어를 감소시킨다.[105]
척추동물은 피부, 입, 위, 눈, 면역 체계 등 다양한 방어 기제를 가지고 있다. 곤충은 기생충을 막기 위해 둥지를 적응시키기도 하며, 식물은 화학 물질을 통해 방어한다.
3. 1. 피부

기생충 침입에 대한 1차 방어선은 피부이다.[106] 피부는 물리적 장벽으로, 죽은 세포층으로 구성되어 있다. 이 죽은 세포에는 케라틴이라는 단백질이 포함되어 피부를 질기고 방수 상태로 만든다. 대부분의 미생물은 생존을 위해 수분이 많은 환경이 필요하므로, 건조한 피부는 생물의 침입과 대량 서식을 막는다. 또한 인간의 피부는 대부분의 미생물에 독성인 피지를 분비한다. 파충류, 조류, 포유류의 질기고 건조하며 방수가 되는 피부는 침입하는 미생물이 몸속으로 들어오는 것을 막는 물리적 장벽 역할을 한다. 반면, 흡충 같은 더 큰 기생충은 물에 들어갈 때 숙주를 찾기 위해 피부에서 생성되는 화학 물질을 감지한다.[106]
3. 2. 입
입은 외래 생물이 입을 통해 침입하는 것을 막는 침을 포함하고 있다. 또한 입안에는 눈물과 타액에서 발견되는 라이소자임이라는 효소가 포함되어 있는데, 이 효소는 미생물이 침입했을 때 세포벽을 파괴한다.[20]
3. 3. 위
위는 생물이 입을 통과한 후 만나는 다음 방어선이다. 위는 위장관에서 체내로 침입하려는 대부분의 미생물을 죽이는 데 도움이 되는 pH 2의 염산 및 위산을 포함하고 있다.[106]
3. 4. 눈
눈꺼풀의 속눈썹은 눈에 들어오는 미생물의 침입을 막는 첫 번째 방어선 역할을 한다. 그럼에도 미생물이 눈 안으로 들어가면, 눈물에 포함된 라이소자임이라는 효소가 침입한 미생물을 제거하는 기능을 한다.[20]
3. 5. 면역 체계
기생충이 체내에 들어오면 면역 체계가 주요 방어 역할을 한다. 면역계는 혈청 단백질과 패턴 인식 수용체(PRR)를 포함한 다양한 분자 결합으로 구성된다.[107] PRR은 T세포, B 세포 등 적응 면역계의 림프구를 활성화시키고, 림프구는 가변 수용체를 통해 기생충을 인식한다.[107]
파충류, 조류, 포유류의 질기고 건조하며 방수가 되는 피부는 기생충의 침입을 막는 물리적 장벽 역할을 한다.[106] 인간의 피부는 대부분의 미생물에 독성인 피지를 분비한다.[106] 흡충과 같은 더 큰 기생충은 물에 들어갈 때 피부에서 생성되는 화학 물질을 감지하여 숙주를 찾는다. 척추동물의 침과 눈물에는 라이소자임이 함유되어 있어 침입하는 박테리아의 세포벽을 분해한다.[106] 입을 통과한 미생물은 염산을 가진 위에 의해 방어된다.[106] 일부 장내 기생충은 두껍고 질긴 외부 코팅을 가지고 있어 소화되지 않고 위를 통과하여 장에서 다음 생애 단계를 시작할 수 있다. 체내에 들어간 기생충은 면역 체계의 혈청 단백질과 패턴 인식 수용체를 극복해야 하며, 이는 적응 면역 체계의 림프구를 유발한다.[107]
3. 6. 동물의 추가적인 방어
영양적으로 전파되는 기생충은 숙주에게 먹힘으로써 전파된다. 여기에는 흡충류(주혈흡충류 제외), 촌충, 극두동물, 설충, 많은 선충류, 그리고 ''톡소포자충''과 같은 많은 원생동물이 포함된다.[20] 이들은 두 종 이상의 숙주를 포함하는 복잡한 생활사를 가지고 있다. 유충 단계에서 중간 숙주를 감염시키고 종종 중간 숙주 내에서 낭포를 형성한다. 중간 숙주 동물이 포식자, 즉 최종 숙주에게 먹히면, 기생충은 소화 과정을 거쳐 살아남아 성충으로 성숙하며, 일부는 장내 기생충으로 산다. 많은 영양적으로 전파되는 기생충들은 중간 숙주의 행동을 수정하여 포식자에게 잡아먹힐 가능성을 높인다. 직접 전파되는 기생충과 마찬가지로, 영양적으로 전파되는 기생충의 숙주 개체 간 분포는 집합적이다.[20] 여러 기생충에 의한 중복 감염이 흔하다.[24] 예외적으로 기생충의 전체 생물학적 생활사가 단일 일차 숙주 내에서 발생하는 자가 감염은 ''분선충''과 같은 선충류에서 때때로 발생할 수 있다.[25]
극상기생은 기생충 안에서 사는 원생동물과 같이 다른 기생충을 먹고 살거나,[34] 숙주가 일반적인 기생충 또는 기생성 천적인 임의적 또는 절대적 기생성 천적의 예와 같다.[20][30] 특히 임의적 기생성 천적 사이에서 2차 기생을 넘어선 기생 수준도 발생한다. 참나무 혹벌 시스템에서는 최대 4단계의 기생이 있을 수 있다.[35]
극상기생은 숙주의 개체수를 조절할 수 있으며, 이러한 목적으로 농업과 어느 정도 의학에서 사용된다. 조절 효과는 CHV1 바이러스가 밤나무 궤양병인 ''Cryphonectria parasitica''가 미국 밤나무에 주는 피해를 조절하는 방식과 박테리오파지가 세균 감염을 제한하는 방식에서 볼 수 있다. 연구는 거의 이루어지지 않았지만, 대부분의 병원성 미세 기생충은 농업과 의학 모두에서 널리 유용할 수 있는 극상기생을 가지고 있을 가능성이 높다.[36]
3. 7. 곤충의 방어
곤충은 종종 기생충을 막기 위해 둥지를 적응시킨다. 예를 들어, 말벌의 일종인 ''Polistes canadensis''는 좀나방 감염을 피하기 위해 여러 개의 벌집판에 둥지를 짓는다.[108] 좀나방은 말벌 둥지에 알을 낳고, 부화한 유충은 말벌 번데기를 먹는다. 다 자란 말벌은 세포 가장자리를 갉아먹고 구강 분비물을 발라 나방 알과 유충을 제거하려 한다.[108]
3. 8. 식물의 방어
식물은 기생충의 공격에 반응하여 폴리페놀 산화 효소를 만드는 등 다양한 화학 방어 물질을 생성하거나, 자스몬산 불감성(JA) 및 살리실산(SA) 신호 전달 경로를 제어한다.[109][110] 이러한 생화학적 경로는 서로 다른 공격에 의해 활성화되며, 긍정적이거나 부정적으로 상호 작용할 수 있다. 식물은 보통 특이적 반응 또는 비특이적 반응을 보인다.[110][111]
특이적 반응은 식물 세포 수용체가 기생충을 인식하면서 시작된다. 이는 강력하지만 국소적인 반응으로 이어진다. 방어 화학 물질은 기생충이 감지된 주변 영역에서 생성되어 확산을 차단하고, 필요하지 않은 곳에서 방어 물질을 만드는 낭비를 막는다.[111]
비특이적 방어 반응은 전신적이다. 즉, 반응이 식물의 특정 영역에 국한되지 않고 식물 전체로 퍼져 에너지 소모가 많다. 하지만, 이러한 반응은 광범위한 기생충에 효과적이다.[111] 예를 들어, 나비목 애벌레에게 손상을 입으면, 옥수수와 목화를 포함한 식물의 잎은 공격을 받고 있음을 알리는 테르펜과 같은 휘발성 화학 물질의 양을 증가시킨다. 이는 기생 말벌을 유인하여 애벌레를 공격하게 한다.[112]
4. 기생의 진화
기생은 다양한 동물, 식물, 균류에서 독립적으로 여러 번 진화해왔다. 모든 동물의 절반 정도는 일생에서 적어도 한 번은 기생 단계를 거치며, 거의 모든 자유생활동물은 하나 이상의 기생 생물 군의 숙주 역할을 한다.[152]
기생충은 숙주의 방어에 대응하여 진화하며, 때로는 특정 숙주 종에 특이적으로 나타나기도 한다. 하지만 숙주 종이 멸종하면 이러한 특이성은 비효율적일 수 있으므로, 많은 기생충은 여러 숙주를 감염시킬 수 있는 능력을 가진다.
숙주 역시 기생충의 공격에 대응하여 방어 기제를 진화시킨다. 기생충은 세대 기간이 짧아 숙주보다 진화 속도가 빠르기 때문에, 진화 경쟁에서 유리한 위치를 차지한다. 일반적으로 기생충은 숙주에게 해를 덜 끼치는 방향으로 진화하며, 숙주는 기생충의 존재에 적응하는 방향으로 진화한다. 예를 들어, 기생충 감염은 동물의 면역 질환을 줄이는 데 도움을 줄 수 있다. 심지어 일부 선충은 볼바키아 균 감염 없이는 생존할 수 없다.
기생충 간의 경쟁은 더 빠른 번식을 유도하며, 이는 더 치명적인 기생충의 출현으로 이어질 수 있다. 숙주의 죽음을 통해 다른 숙주로 이동하는 기생충은 더욱 치명적으로 진화하며, 숙주의 행동을 변화시켜 포식자에게 더 취약하게 만들기도 한다.
기생충과 숙주의 공진화 역사는 숙주 분류군 간의 관계를 밝히는 데 도움을 줄 수 있다. 예를 들어, 플라밍고가 황새보다는 오리와 더 가까운 관계라는 사실은 이들이 공유하는 기생충을 통해 알 수 있다.
기생은 수컷 공작의 깃털이나 수컷 사자의 갈기와 같은 동물 수컷의 이차 성징 진화에 대한 설명의 일부이기도 하다. 암컷 숙주는 기생충과 질병에 대한 저항성을 나타내는 이러한 특성을 가진 수컷을 선택하여 번식한다.
도둑기생은 기생 생물이 숙주가 모은 먹이를 훔치는 현상이다. 이는 같은 종 내, 또는 가까운 친척 관계의 종 사이에서 발생한다. 예를 들어, 뻐꾸기벌은 다른 벌의 둥지에 알을 낳는다. 슴새와 같은 일부 조류는 다른 바닷새의 먹이를 빼앗는 데 특화되어 있다.[51][52]
기생은 생물 간 상호 작용의 한 형태이며, "여러 종의 생물이 상호 작용을 하면서 같은 장소에서 생활하는" 공생에 포함된다. 기생은 다음과 같이 정의할 수 있다.
- A가 B가 얻었거나 얻을 영양 섭취원을 획득한다.
- A가 B의 체내, 체표면, 또는 가까운 위치에 있다.
- 이 관계가 일정 기간 지속된다.
- B는 A의 존재로 인해 불이익을 받는다.
이러한 관계에서 A는 B에 '기생'하며, B는 A의 '숙주'가 된다.
회충이나 이는 사람에게 기생하는 대표적인 예시이다. 반면, 모기나 파리는 사람의 피를 빨지만, 짧은 시간 동안만 접촉하므로 기생자로 분류되지 않는다.
충영을 형성하는 곤충은 기생자로 간주되지만, 잎을 먹는 애벌레는 일반적으로 식식자로 분류된다. 포식기생은 숙주를 죽이는 기생 형태로, 벌이나 파리에서 흔히 나타난다. 사마귀붙이나 알노린재처럼 알을 먹는 경우도 기생이라고 불리지만, 이는 단순한 알 포식이다.
기생 생물은 체 기능의 많은 부분을 기생 대상에 의존하기 때문에, 특히 내부 기생 종은 신체 구조가 퇴화하는 경향이 있다. 배좀벌레나 오구동물은 극단적인 퇴행적 특수화를 보인 예시이다. 반면, 기생 생물은 숙주의 방어 기능을 피하기 위한 생리 기능이나 복잡한 생활환을 진화시키기도 한다. 척추동물은 변태 기능이 발달하지 않아 내부 기생 사례가 없다.
심해 아귀류의 수컷은 암컷에 비해 매우 작고, 암컷의 몸에 붙어 기생 생활을 한다. 이는 심해 환경에서 번식 기회를 확보하기 위한 적응이다. 미츠쿠리에나가지초롱아귀와 같은 종은 수컷의 기관이 퇴화하여 암컷과 거의 동화된다. 이러한 수컷을 왜소웅이라고 한다.
태생 동물에서 태아는 어머니에게서 영양을 공급받으므로, 기생 관계라고 볼 수 있다. 종자식물의 본체와 배우체 사이에도 비슷한 관계가 성립한다.
촌충은 장내에 머물면서 영양을 빼앗지만, 면역계를 자극하여 건강 유지에 도움을 줄 수 있다는 연구 결과도 있다.
진딧물의 구기나 바이러스의 유전자 재작성 능력은 유전자 조작에 응용되기도 한다.
고착성 동물 중에는 뿌리 모양의 구조를 동물체 내에 침투시켜 고정하는 경우가 있는데, 이는 기생 생활로 이어지는 경로 중 하나로 여겨진다.
부생 식물은 과거에는 사물기생을 하는 것으로 여겨졌으나, 현재는 '''균종속영양 식물'''이라고 불린다. 이들은 균근균에게서 영양을 얻으며, 난초과 식물에서 다양한 형태로 나타난다.
균류에서 살아있는 생물로부터 직접 영양을 얻는 것은 기생성 균류라고 불린다. 느타리버섯과 같은 일부 버섯은 선충 포식능을 가지며, 털곰팡이목의 포식성 균류는 소동물을 포획하여 영양을 흡수한다.
백선균 (무좀)은 각질을 침식하는 기생균이지만, 부생균과의 구별이 모호한 경우도 있다.
4. 1. 공진화
기생충과 숙주는 서로에게 영향을 주며 함께 진화한다. 예를 들어 원숭이 포말상 바이러스(SFV)와 영장류 숙주는 공진화의 대표적인 예시이다. 한 연구에서 아프리카와 아시아 영장류의 SFV 효소와 미토콘드리아 시토크롬 산화 효소 서브 유닛 II의 계통 발생을 비교했을 때, 놀랍게도 계통 발생에서 분기 순서와 횟수가 일치했다.[94] 이는 원숭이 포말상 바이러스가 최소 30만 년 동안 전 세계 영장류와 함께 진화해 왔음을 보여준다.
기생충과 숙주의 공진화 역사를 추정하면 숙주 분류군 간의 관련성을 밝힐 수 있다. 예를 들어, 홍학이 황새와 더 가까운지, 아니면 오리와 더 가까운지에 대한 논쟁이 있었다. 홍학이 오리, 거위와 같은 기생충을 공유한다는 사실은 이들이 황새보다 서로 더 가깝다는 것을 시사했다. 그러나 기생충 종의 중복이나 멸종과 같은 진화적 사건은 숙주와 기생충 계통수 간의 유사성을 약화시키기도 한다. 홍학의 경우, 논병아리와 유사한 이가 있는데, 이는 홍학과 논병아리가 공통 조상을 가지고 있으며 이들 그룹에서 조류와 이의 공생 진화를 시사한다. 이후 홍학의 이는 숙주 전환을 통해 오리로 옮겨갔다.[95]
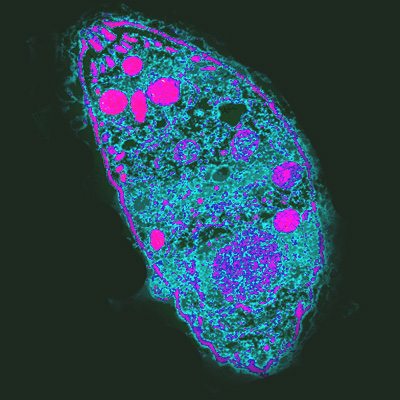
기생충은 동소적 종(같은 지역) 숙주를 더 효과적으로 감염시킨다. 이는 호수 달팽이를 감염시키는 흡충류에서 확인되었다.[96] 이는 종 간의 상호작용이 지속적인 자연 선택으로 이어져 공진화를 유도한다는 레드 퀸 가설과 일치한다. 기생충은 지역적으로 흔한 숙주의 표현형을 추적하므로, 다른 지역의 숙주에는 덜 감염된다.[96]
4. 2. 붉은 여왕 가설
기생충은 같은 지리적 영역(같은 곳)에 존재하는 숙주를 더 효과적으로 감염시킨다. 이 현상은 종간의 상호작용(기생충과 숙주 같은)이 적응과 대응을 위한 지속적인 자연 선택을 야기한다고 주장하는 “붉은 여왕 가설”을 뒷받침한다.[96] 기생충은 지역적으로 일반적인 숙주의 표현형을 쫓아가므로, 다른 지역의 숙주에 대해서는 전염성이 덜하다.[96]2000년에 출판된 실험은 뉴질랜드의 Ianthe 호수와 Poerua 호수에서 두 개의 서로 다른 달팽이 집단의 분석을 논의한다. 집단은 같은 호수에서 채취한 2개의 순수한 기생충(이생적 흡충)에 노출되었다. 실험에서 달팽이는 같은 지역의 기생충, 다른 지역의 기생충, 그리고 두 기생충을 혼합한 것에 감염되었다. 결과는 기생충이 자신의 지역과 다른 지역의 달팽이보다 같은 지역의 달팽이를 감염시키는데 더욱 효과적이었음을 시사한다. 다른 지역의 달팽이 역시 기생충에 감염되었지만, 전염성은 같은 지역의 달팽이와 비교하면 훨씬 적었다. 따라서, 기생충은 그 지역의 달팽이 집단 감염에 더 적합한 것으로 나타났다.
4. 3. 행동 변화
일부 기생충은 숙주 간의 전파를 늘리기 위해 숙주의 행동을 수정하는데, 이는 종종 포식자와 피식자의 관계와 관련이 있다(기생충에 의한 영양 단계 전파 증가).[97]- 캘리포니아 해안 염습지에 사는 흡충류 ''Euhaplorchis californiensis''는 숙주인 킬리피쉬가 포식자를 피하는 능력을 감소시킨다.[97] 이 기생충은 백로에서 성숙하며, 백로는 감염되지 않은 물고기보다 감염된 킬리피쉬를 먹을 가능성이 더 높다.
- 고양이에서 성숙하지만 다른 많은 포유류도 옮길 수 있는 원생동물 ''톡소포자충''에 감염된 시궁쥐는 고양이 냄새에 이끌리지만, 감염되지 않은 쥐는 고양이 냄새를 피한다. 이는 톡소포자충이 고양이 숙주로 전파될 가능성을 높인다.[98]
- 말라리아 기생충은 인간 숙주의 피부 냄새를 변형시켜 모기에 대한 매력을 증가시키고, 따라서 기생충이 전파될 기회를 높인다.[33]
- 거미 ''Cyclosa argenteoalba''는 기생충 벌레 유충에 의해 거미집을 짓는 행동이 바뀐다. 기생충이 부착되면 정상적인 끈적한 나선형 거미집 대신 단순화된 거미집을 만든다. 이 행동은 기생충이 거미에 오래 남아있을수록 더 두드러진다.[99]
기생자가 숙주에게 특수한 행동을 취하게 하는 예는 다음과 같다.[150]
- 흡충류 백록클로리디움(Leucochloridium)의 유생은 논우렁이에 기생한다. 유생이 성숙하면 논우렁이 눈으로 이동하여 눈 자루를 크게 부풀게 하고, 유생의 줄무늬가 화려하게 움직이는 것처럼 보이게 한다. 논우렁이는 평소 그늘에 숨지만, 이때는 잎 표면에 나타나 새의 눈에 띄게 된다. 새가 논우렁이 눈을 쪼아 먹으면 유충이 튀어나와 새의 체내로 들어간다.
- 사다리벌레에 기생된 게 등 갑각류 수컷은 암컷처럼 행동한다. 흉부에 굽어진 복부가 넓어지고 알을 품는 암컷과 같은 형태가 된다. 사다리벌레는 게가 포란할 때 알이 들어갈 장소에 발달하여, 게가 사다리벌레를 알 덩어리로 착각하고 지키는 행동을 하게 만든다.
- 말벌레는 숙주를 물가로 유도하여 물속에서 나온다. 숙주의 생식 기능을 상실시키고, 뇌에 영향을 주는 단백질을 주입하며, 물가로 유도할 때는 기생한 사마귀의 편광을 식별하는 시각을 이용한다.[150][151]
5. 기생의 생태
기생은 진화 생태학의 주요 측면이다. 거의 모든 자유 생활 동물은 적어도 한 종 이상의 기생충을 가지고 있다. 척추동물은 75,000~300,000종의 헬민스와 수많은 기생 미생물의 숙주이며, 포유류는 평균적으로 4종의 선충, 2종의 흡충, 2종의 촌충을 숙주로 한다. 인간은 342종의 헬민스 기생충과 70종의 원생동물 기생충을 가지고 있다.[113] 전체 종의 약 40%가 기생충이며, 먹이 그물에서 약 4분의 3의 연결은 기생충을 포함한다.[81]
기생충과 기생충의 진화는 21세기까지 주로 기생충학자들에 의해 연구되었으며, 생태학자나 진화 생물학자보다는 의학이 지배적인 과학이었다. 기생충-숙주 상호작용은 생태적이고 진화에 중요한 요소였지만, 기생충학의 역사는 진화 생태학자 로버트 풀린이 "기생충학자들에 의한 기생의 장악"으로 표현한 상황을 초래하여 생태학자들이 이 분야를 무시하게 만들었다. 풀린은 기생충이 "자연 선택의 편재하는 요인"이며 진화와 생태학에서 중요한 힘을 가지고 있다는 점에서 이는 "불행한 일"이라고 생각했다.
복잡한 관계로 인해 기생충을 먹이 그물에 배치하기는 어렵다. 여러 숙주를 가진 흡충은 동시에 먹이 그물의 여러 위치를 차지하고 에너지 흐름의 루프를 설정하여 분석을 혼란스럽게 한다. 또한 거의 모든 동물이 여러 기생충을 가지고 있기 때문에, 기생충은 모든 먹이 그물의 최상위 수준을 차지한다.[113]
기생충은 비토착 종의 확산에 영향을 줄 수 있다. 예를 들어, 침입성 녹색 게는 동부 대서양 해안의 토착 흡충류의 영향을 최소한으로 받아 토착 게와의 경쟁에서 우위를 점한다.[114]
생태적 기생충학은 드라쿤쿨루스증 퇴치 캠페인과 같이 통제 시도에 중요할 수 있다. 기생충이 4개국을 제외한 모든 국가에서 근절되었음에도 불구하고, 벌레는 개를 감염시키기 전에 개구리를 중간 숙주로 사용하기 시작하여 이러한 관계가 더 잘 이해되었다면 통제가 더 어려워졌을 것이다.[115]
기생충은 일반적으로 해로운 존재로 여겨지지만, 모든 기생충을 없애는 것이 유익하지는 않다. 기생충은 생물 다양성의 최소 절반을 차지하며, 중요한 생태학적 역할을 수행한다. 기생충이 없다면 생물은 무성 생식을 선호하여 유성 생식으로 인한 특성의 다양성이 감소할 수 있다.[116] 기생충은 종 간의 유전 물질 전달 기회를 제공하여 진화적 변화를 촉진한다.[117] 많은 기생충은 생활 주기를 완료하기 위해 여러 종의 숙주가 필요하며, 포식-피식 관계 또는 기타 안정적인 생태적 상호 작용에 의존하여 한 숙주에서 다른 숙주로 이동한다. 따라서 기생충의 존재는 생태계가 건강하다는 것을 나타낸다.[118]
캘리포니아 콘도르의 외기생충인 캘리포니아 콘도르 이(''Colpocephalum californici'')는 캘리포니아 콘도르를 구하기 위한 사육 프로그램 과정에서 의도적으로 제거되어 멸종되었다.[119]
기생충은 종종 먹이 사슬 묘사에서 생략되지만, 일반적으로 최상위에 위치한다. 기생충은 핵심종과 같이 작용하여 우세한 경쟁자의 지배력을 감소시키고 경쟁 종이 공존할 수 있게 한다.[113][120][121]
5. 1. 정량 생태
기생충은 숙주 개체군 내에서 집괴 분포를 보이는 경향이 있는데, 이는 대부분의 숙주가 적은 수의 기생충을 가지고, 소수의 숙주가 많은 수의 기생충을 가진다는 것을 의미한다.[122] 이러한 분포는 생물학자들이 일반적으로 사용하는 모수 통계를 무효화하여 기생충 생태학 연구에 어려움을 야기한다.[122] 따라서 로그 변환 후 모수 검정을 적용하거나 비모수 통계를 사용하는 것이 권장되기도 하지만, 이 역시 또 다른 문제를 야기할 수 있어 현대 정량 기생충학은 더 발전된 생물 통계적 방법에 기반한다.[122]5. 2. 생태 다양성
기생충은 숙주를 서식지로 이용하여 다양한 생태적 지위를 차지한다. 서로 다른 규모의 숙주에서 기생충의 집합체를 설명하기 위해 다음과 같은 용어들이 계층적으로 사용된다.
기생충의 생태 다양성은 자유생활생물의 그것과는 크게 다르다. 자유생활생물에 있어서 생태 다양성은 섬 생물 지리학의 로버트 맥아더(Robert MacArthur)와 E. O. 윌슨(E. O. Wilson)의 이론, 제러드 다이아몬드(Jared Diamond)의 조립 규칙, 그리고 최근에는 생물 다양성과 생물 지리학의 스티븐 허벨(Stephen Hubbell)의 통합 중립 이론의 null 모델을 포함하여 많은 강력한 개념적 틀을 제공한다. 이러한 틀들은 기생충을 위해서는 충분히 개발되어 있지 않고, 기생충에게 자유생활생물의 모델은 많은 점에서 적합하지 않다. 예를 들어, 섬 생물 지리학은 일반적으로 본토를 참조하여 서식 구역 사이의 고정된 공간적인 관계를 기반으로 한다. 기생충은 동적 공간 관계를 갖는 움직이는 서식지인 숙주에 살고 있다. 숙주 집단이 아닌 진정한 ‘본토’는 없기에 숙주 내에서 공동체를 구성하는 기생충들은 metacommunities이다.
그럼에도 불구하고, 서로 다른 기생충 집합체의 종류는 단일 숙주나 집단 내에서 인식되고 있으며, 자유생활생물에서 관찰된 패턴의 대부분은 기생충 집합체에서도 나타난다. 이들 중 가장 눈에 띄는 것은 상호 고립적 연속체이다. 이것은 기생충의 집합체가 고립계에 있어서 생태적 지위가 포화되고 이종 간의 경쟁이 만연하는 공동체 또는 생태적 지위에 여유가 있고 사회구조형성에 있어서 확률 요인만큼 이종 간의 상호작용이 중요하지 않은 공동체에서 나타나는 연속변이를 따라 발생한다고 주장한다. 이것이 정말 이런 것인지 또는 이러한 패턴이 단순히 존재하는 종의 분포를 반영하고 있는지 여부는 아직 확립되어 있지 않다.
기생충은 비토착 종의 확산에 역할을 할 수 있다. 예를 들어, 침입성 녹색 게는 동부 대서양 해안의 토착 흡충류의 영향을 최소한으로 받는다. 이는 대서양 바위 게와 조나 게와 같은 토착 게와의 경쟁에서 우위를 점하도록 돕는다.[114]
생태적 기생충학은 드라쿤쿨루스증 퇴치 캠페인과 같이 통제 시도에 중요할 수 있다. 기생충이 4개국을 제외한 모든 국가에서 근절되었음에도 불구하고, 벌레는 개를 감염시키기 전에 개구리를 중간 숙주로 사용하기 시작하여 이러한 관계가 더 잘 이해되었다면 통제가 더 어려워졌다.[115]
5. 3. 생태계 내 역할
기생충은 종종 먹이 사슬에서 생략되지만, 일반적으로 최상위 포식자 위치를 차지한다.[113] 이들은 우점종의 경쟁을 억제하여 다른 종들이 공존할 수 있도록 돕는 핵심종 역할을 수행하기도 한다.[113][120][121]대부분의 기생충은 생활사를 완료하기 위해 여러 숙주를 필요로 하며, 포식-피식 관계나 다른 안정적인 생태적 상호작용에 의존한다.[118] 이러한 점에서 생태계 내 기생충의 존재는 그 체제의 건강 상태를 반영한다고 볼 수 있다.
예를 들어, 침입종인 녹색 게는 동부 대서양 해안의 토착 흡충류의 영향을 적게 받아, 토착 게들과의 경쟁에서 우위를 점할 수 있다.[114]
기생충은 숙주의 행동을 변화시켜 다른 숙주로의 전파를 용이하게 하기도 한다. 예를 들어, 캘리포니아 해안 염습지의 흡충류인 ''Euhaplorchis californiensis''는 숙주인 킬리피쉬가 포식자를 피하는 능력을 감소시킨다.[97] 이 기생충은 백로에서 성숙하는데, 백로는 감염된 킬리피쉬를 더 잘 잡아먹는다.
기생충은 일반적으로 해로운 존재로 인식되지만, 생태계에서 중요한 역할을 수행한다. 모든 기생충을 제거하는 것은 생물 다양성을 감소시키고, 유성 생식으로 인한 특성의 다양성을 줄일 수 있다.[116] 또한, 기생충은 종 간의 유전 물질 전달 기회를 제공하여 진화적 변화를 촉진한다.[117]
6. 기생충 전파
기생충은 다양한 방법으로 숙주를 감염시킨다. 예를 들어, 아칸트아메바는 적절한 환경이 되면 체내로 들어가고, 분선충은 인간이 맨발로 감염된 땅을 걸어 다닐 때 체내로 들어온다. 사일열원충은 말라리아를 퍼뜨리기 위해 모기를 숙주로 이용하고, 로아사상충은 사슴파리를 통해 숙주 내로 들어간다.
기생충은 살아있는 생물을 서식지로 삼기 때문에 자유생활생물에게는 없는 문제에 직면한다. 기생충이 살아남을 수 있는 유일한 서식지인 숙주가 기생충을 피하거나, 격퇴하거나, 제거하려고 하기 때문이다. 이에 기생충은 정착, 점유, 전파 등의 과정을 통해 다른 숙주를 찾는 전략을 사용한다.
일부 내부 기생충은 섭취되어야 하는 것들과 달리 숙주의 외부 표면을 뚫고 감염시키기도 한다. 숙주 내부에 들어온 내부 기생충은 다른 숙주를 감염시키기 위해 자손을 외부로 내보내야 한다.
6. 1. 직접 접촉
단생흡충과 같은 몇몇 외부 기생충은 숙주와의 직접적인 접촉에 의존한다. 외부 기생 절지동물은 숙주 간의 접촉(대부분의 이)에 의존하거나, 숙주에서 살아남기 위해 알을 낳거나(예: 벼룩), 숙주를 만나기 위한 외부 환경을 기다린다(예: 진드기). 일부 수생 거머리는 움직임을 감지하여 숙주의 위치를 파악하고 특정한 온도나 화학 신호가 존재할 때만 달라붙는다.[75]6. 2. 분변-경구 경로
이질 아메바와 같은 많은 내부 기생충은 자손이 숙주의 배설물과 함께 흘러나갈 수 있도록 숙주의 위장관에 존재한다. 구두동물, 촌충, 흡충 등이 이 방법을 사용한다.[1]6. 3. 매개체
사일열원충은 말라리아를 퍼뜨리기 위해 숙주로 모기를 이용하고, 로아사상충은 숙주 내로 들어가기 위해 사슴파리를 이용한다.[74] 말라리아 기생충과 트리파노소마 등의 내부 기생 원충은 무는 벌레나 매개체들에 의해 새로운 숙주에게 전파된다.[28]라임병과 재귀열의 원인인 ''보렐리아''는 사슴과 같은 동물에 있는 질병의 저장소에서 ''참진드기''와 같은 매개체를 통해 전파된다.[76]
6. 4. 포식-피식 관계
내부 기생충의 유충은 종종 혈액 또는 위장관 이외의 숙주 내부를 감염시킨다. 그런 경우 대부분은 생존과 번식을 위해 기생충의 생애주기에 있어서 숙주가 다음 숙주에게 잡아먹히는 것을 필요로 한다. 위의 전략은 구두동물, 촌충, 흡충 그리고 기생 회충의 유충 단계에 따라 다양하게 사용되고 있다.[1]일부 기생충은 보통 피식자와 포식자 관계에 놓여있는 숙주간의 전염을 증가시키기 위해 숙주의 행동을 변화시킨다. 예를 들어, 캘리포니아의 갯벌에서 ''Euhaplorchis californiensis'' 흡충은 포식자를 피하기 위한 숙주의 능력을 저하시킨다. 이 기생충은 감염되지 않은 것보다 감염된 물고기를 먹을 확률이 더 높은 왜가리 안에서 성숙한다. 또 다른 예로는 고양이 안에서 성숙하지만 다른 포유류로 전달될 수 있는 톡소플라즈마원충이 있다. 감염되지 않은 쥐는 고양이의 냄새를 피하지만, 톡소플라즈마에 감염된 쥐는 고양잇과 숙주에게로의 전염을 증가시킬 수 있는 냄새에 이끌린다.[1]
7. 기생의 가치
기생충은 일반적으로 해롭다고 생각되지만, 모든 기생충을 근절하는 것이 반드시 유익한 것은 아니다. 기생충은 생물 다양성의 절반 이상을 차지하며, 먹이 사슬에서 핵심종과 같이 작용하여 우세한 경쟁자의 지배력을 감소시키고 다른 종들이 공존할 수 있게 돕는 중요한 역할을 수행한다.[113][120][121]
또한, 기생충이 숙주에게 약간의 이익을 가져다주는 경우도 있다. 예를 들어, 인간의 촌충은 장내에 머물면서 영양을 빼앗지만, 동시에 면역계를 자극하여 건강 유지에 도움을 줄 수 있다는 연구 결과도 있어 기생인지 공생인지에 대한 논쟁의 대상이 되기도 한다.
캘리포니아 콘도르를 구하기 위한 프로그램에서 콘도르에만 서식하는 캘리포니아 콘도르 이를 제거한 결과, 콘도르는 구출되었지만 기생충은 멸종된 사례는 보존 문제에 대한 시사점을 제공한다.[119]
7. 1. 유전적 다양성 증가
기생충은 일반적으로 유해하다고 생각되지만, 모든 기생충 근절이 절대 유익하지는 않다. 기생충은 생명 다양성의 절반 이상을 차지하며, 먹이를 약화시키는 중요한 생태학적 역할을 수행한다. 생태계와 그 구성요소들은 각각 적응하는 데 시간이 필요하다. 기생충이 존재하지 않는다면 생명체들은 성적 이형 요소를 감소시키며 무성 생식을 할지도 모른다. 기생충은 종간의 유전적 물질 이동을 위한 기회를 제공한다. 드물지만 중요하게도, 이는 기생충이 아니라면 발생하지 않거나 발생한다 해도 오랜 시간이 걸리는 진화를 촉진시킨다.[1]7. 2. 생태계 건강 지표
기생충은 일반적으로 해로운 존재로 여겨지지만, 모든 기생충을 없애는 것이 유익한 것은 아니다. 기생충은 생물 다양성의 최소 절반을 차지하며, 먹이를 약화시키는 중요한 생태학적 역할을 수행한다. 기생충이 없다면 생물은 무성 생식을 선호하여 유성 생식으로 인한 특성의 다양성이 감소할 수 있다.[116] 기생충은 종 간의 유전 물질 전달 기회를 제공하여 진화적 변화를 촉진한다.[117] 많은 기생충은 생활 주기를 완료하기 위해 여러 다른 종의 숙주가 필요하며, 포식-피식 관계 또는 기타 안정적인 생태적 상호 작용에 의존하여 한 숙주에서 다른 숙주로 이동한다. 따라서 기생충의 존재는 생태계가 건강하다는 것을 나타낸다.[118]
8. 한국의 기생충 연구 및 관리
주어진 원본 소스에는 한국의 기생충 연구 및 관리에 대한 구체적인 내용이 없으므로, 주어진 정보와 요약만으로는 이 섹션의 내용을 작성할 수 없다. 요약에 따르면 한국은 국가적인 기생충 관리 사업을 통해 기생충 질환 유병률을 크게 감소시켰다고는 하나, 원본 소스에는 관련 내용이 전혀 없어 내용을 추가할 수 없다.
8. 1. 역사
인간 기생충에는 회충, 기니벌레, 요충, 촌충 등이 있으며, 기원전 3000년 이후 이집트 파피루스 기록에 언급되어 있다. 에버스 파피루스는 구충을 묘사하고 있다. 고대 그리스에서는 방광충을 포함한 기생충이 히포크라테스 전집에 묘사되어 있으며, 희극 작가 아리스토파네스는 촌충을 "우박"이라고 불렀다. 로마의 의사 켈수스와 갈레노스는 회충 ''Ascaris lumbricoides''와 ''Enterobius vermicularis''를 기록했다.[123]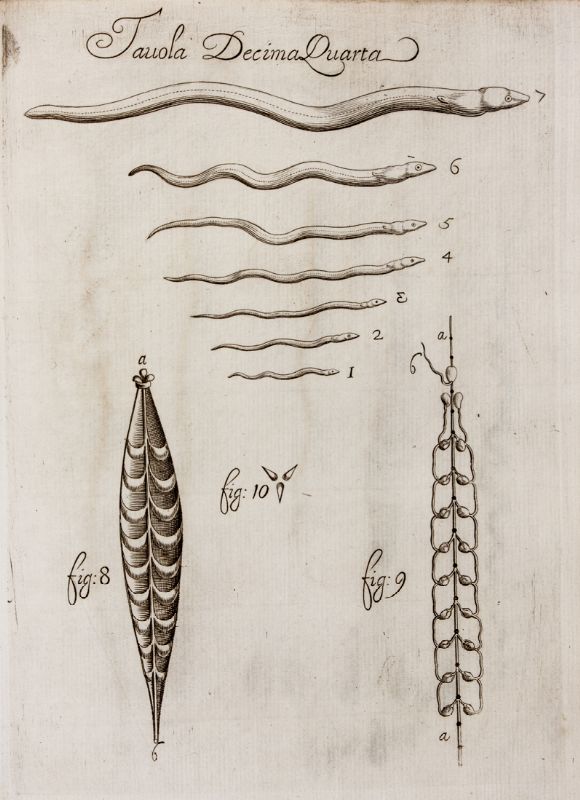
1025년에 완성된 아비센나의 저서 ''의학 정전''에는 둥근벌레, 실벌레, 기니벌레, 촌충을 포함한 인간과 동물의 기생충이 기록되어 있다.[123]
1397년 저서 ''Traité de l'état, science et pratique de l'art de la Bergerie'' (양치기 기술의 상태, 과학 및 실천에 대한 설명)에서 Jehan de Brie|장 드 브리프랑스어는 흡충류 내부 기생충인 양 간 플루크 ''간질''(Fasciola hepatica)에 대한 최초의 설명을 기록했다.[128][124]
근대 초기 시대에 프란체스코 레디는 1668년 저서 《곤충의 발생에 관한 경험》(''Esperienze Intorno alla Generazione degl'Insetti'')에서 외부 및 내부 기생충을 명확히 묘사하며, 진드기, 사슴 코파리의 유충, 양 간흡충을 예시로 들었다.[125] 레디는 기생충이 알에서 발생하며, 이는 자연 발생설에 반하는 것이라고 언급했다.[126] 1684년 저서 《살아있는 동물 속에 존재하는 살아있는 동물에 대한 관찰》(''Osservazioni intorno agli animali viventi che si trovano negli animali viventi'')에서 레디는 사람 회충을 포함하여 100종 이상의 기생충을 묘사하고 그림으로 나타냈으며, 이는 회충증을 유발한다.[125] 레디는 개와 양에서 발견되는 ''Echinococcus granulosus''의 낭포를 기생충으로 처음 명명했으며, 1세기 후인 1760년 페터 시몬 팔라스는 이를 촌충의 유충이라고 정확하게 제안했다.[123]
1681년 안토니 판 레이우엔훅은 원생동물 기생충인 ''Giardia lamblia''를 관찰하고 그림으로 묘사했으며, 이를 "자신의 설사"와 연결시켰다. 이것은 현미경으로 관찰된 최초의 인간 원생동물 기생충이었다.[123] 몇 년 후인 1687년, 이탈리아 생물학자 조반니 코시모 보노모와 디아친토 체스토니는 옴이 기생성 진드기인 ''Sarcoptes scabiei''에 의해 발생한다고 묘사하며, 이것을 알려진 현미경적 원인 물질을 가진 인간의 첫 번째 질병으로 기록했다.[127]

현대 기생충학은 19세기에 많은 연구자와 임상가들의 정확한 관찰과 실험을 통해 발전했다.[128] 이 용어는 1870년에 처음 사용되었다.[129] 1828년, 제임스 아너슬리는 아메바증을 설명했는데, 이는 장과 간의 원생동물 감염으로, 병원체인 ''Entamoeba histolytica''는 1873년 프리드리히 뢰슈에 의해 발견되었다. 제임스 파제트는 1835년 인간의 장 선충인 ''Trichinella spiralis''를 발견했다. 제임스 매코넬은 1875년 인간의 간흡충인 ''Clonorchis sinensis''를 기술했다.[123] 앨저넌 토머스와 루돌프 로이카르트는 1881-1883년 실험을 통해 양 간흡충의 생활사를 독립적으로 처음 발견했다.[128] 1877년 패트릭 맨슨은 모기에 의해 전파되는 사상충의 생활사를 발견했다. 맨슨은 더 나아가 말라리아 기생충인 ''Plasmodium''이 모기를 매개체로 한다는 것을 예측하고, 로널드 로스에게 조사를 설득했다. 로스는 1897-1898년에 그 예측이 옳다는 것을 확인했다. 동시에 조반니 바티스타 그라시 등은 ''Anopheles'' 모기에서 말라리아 기생충의 생활사 단계를 기술했다. 로스는 그의 업적으로 인해 논란 속에 1902년 노벨상을 수상한 반면, 그라시는 수상하지 못했다.[123] 1903년, 데이비드 브루스는 아프리카 트리파노소마증의 원생동물 기생충과 체체파리 매개체를 확인했다.[130]
8. 2. 현재 상황
기생충은 숙주를 이용하여 여러 기능을 수행하며, 이러한 기능 상실은 기생충에게 생식에 유리하게 작용할 수 있다. 빈대, 박쥐빈대, 이, 벼룩과 같은 곤충 외부 기생충은 비행 능력을 잃고 숙주에 의존하여 이동한다.[100] 기생충 사이에서 형질 손실은 흔하게 나타난다.[101] 어류의 외부 기생충인 점액포자충 ''Henneguya zschokkei''는 유산소 호흡 능력을 상실한 유일한 동물로 알려져 있으며, 세포에 미토콘드리아가 없다.[102]참조
[1]
논문
Biomass transformation webs provide a unified approach to consumer-resource modelling
[2]
웹사이트
παράσιτος
https://www.perseus.[...]
Liddell, Henry George; Scott, Robert
[3]
웹사이트
παρά
https://www.perseus.[...]
Henry George Liddell, Robert Scott
[4]
웹사이트
σῖτος
https://www.perseus.[...]
Liddell, Henry George; Scott, Robert
[5]
웹사이트
σιτισμός
https://www.perseus.[...]
Liddell, Henry George; Scott, Robert
[6]
서적
Overview of Parasitology
http://parasite.org.[...]
Australian Society of Parasitology and Australian Research Council/National Health and Medical Research Council) Research Network for Parasitology
2010-07
[7]
논문
Ecological and Evolutionary Stabilities of Biotrophism, Necrotrophism, and Saprotrophism
http://pure.iiasa.ac[...]
[8]
논문
Definitions of parasitism, considering its potentially opposing effects at different levels of hierarchical organization
[9]
서적
The Meaning of Human Existence
W. W. Norton & Company
[10]
웹사이트
A Classification of Animal-Parasitic Nematodes
http://plpnemweb.ucd[...]
2016-02-25
[11]
논문
Classification of Human Parasites, Vectors, and Similar Organisms
[12]
서적
Overview of Parasitology
http://parasite.org.[...]
Australian Society of Parasitology and Australian Research Council/National Health and Medical Research Council) Research Network for Parasitology
2010-07
[13]
논문
The mesoparasitic copepod Pennella balaenopterae and its significance as a visible indicator of health status in dolphins (Delphinidae): a review
http://www.oers.ca/j[...]
2018-04-11
[14]
웹사이트
Parasitism {{!}} The Encyclopedia of Ecology and Environmental Management
https://search.credo[...]
Blackwell Science
2018-04-08
[15]
논문
Pugnose eels, ''Simenchelys parasiticus'' (Synaphobranchidae) from the heart of a shortfin mako, ''Isurus oxyrinchus'' (Lamnidae)
https://www.research[...]
[16]
논문
Host vibration—a cue to host location by the parasite, ''Biosteres longicaudatus''
[17]
논문
Multi-cue integration: how female mosquitoes locate a human host
[18]
논문
Host Cues Mediate Growth and Establishment of Oak Mistletoe (Phoradendron leucarpum, Viscaceae), an Aerial Parasitic Plant
[19]
논문
The Many Roads to Parasitism: A Tale of Convergence
https://books.google[...]
Academic Press
[20]
논문
Evolution of parasitism along convergent lines: from ecology to genomics
2015-02
[21]
논문
Trophic strategies, animal diversity and body size
http://homes.msi.ucs[...]
2002
[22]
논문
Infestation of parasitic barnacle Sacculina spp. in commercial marine crabs
2013
[23]
서적
General Parasitology
https://books.google[...]
Elsevier Science
[24]
논문
Concomitant infections, parasites and immune responses
https://researchonli[...]
[25]
웹사이트
Helminth Parasites
http://parasite.org.[...]
Australian Society of Parasitology
2017-10-09
[26]
웹사이트
Pathogenic Parasitic Infections
http://www.peoi.org/[...]
PEOI
2013-07-18
[27]
논문
Lyme disease
2001-07
[28]
논문
Malaria and trypanosome transmission: different parasites, same rules?
[29]
논문
Predation, Herbivory, and Parasitism
https://www.nature.c[...]
2018-02-12
[30]
서적
The Insects: An Outline of Entomology
https://archive.org/[...]
Wiley
2010
[31]
논문
What is a vector?
2017-03
[32]
논문
Networks and the ecology of parasite transmission: A framework for wildlife parasitology
2013-12
[33]
논문
Odours of Plasmodium falciparum-infected participants influence mosquito–host interactions
2017-08
[34]
논문
On Protozoa hyperparasitic in Helminth, with some observations on ''Nosema helminthorum'' Moniez, 1887
1957
[35]
논문
On the Biology of the Inhabitants of Oak Galls of Cynipidae (Hymenoptera) in Britain
https://www.research[...]
1961
[36]
논문
The role of hyperparasitism in microbial pathogen ecology and evolution
2016-01
[37]
논문
Corruption of ant acoustical signals by mimetic social parasites: Maculinea butterflies achieve elevated status in host societies by mimicking the acoustics of queen ants
[38]
논문
Sneaky queens in Melipona bees selectively detect and infiltrate queenless colonies
2013-09
[39]
웹사이트
Social Parasites in the Ant Colony
https://www.antkeepe[...]
Antkeepers
2016-04-04
[40]
논문
Über den Ursprung der dulotischen, parasitischen un myrmekophilen Ameisen
1909
[41]
웹사이트
Social Parasitism in Ants
http://www.nature.co[...]
Nature Education Knowledge
2010-10-29
[42]
논문
Über den Ursprung der dulotischen, parasitischen und myrmekophilen Ameisen
https://archive.org/[...]
1909
[43]
논문
Alternative adaptations, sympatric speciation and the evolution of parasitic, inquiline ants
1991-07
[44]
논문
Parasitic nursing behavior in the wedge-capped capuchin monkey (''Cebus olivaceus'')
[45]
서적
The Cuckoos
Oxford University Press
2005-09-15
[46]
서적
Host–parasite evolution: General principles and avian models
Oxford University Press
1997
[47]
논문
A model system for coevolution: avian brood parasitism
[48]
논문
A novel method of rejection of brood parasitic eggs reduces parasitism intensity in a cowbird host
2013-03
[49]
논문
A parasite in wolf's clothing: hawk mimicry reduces mobbing of cuckoos by hosts
[50]
논문
Spatial variation in egg polymorphism among cuckoo hosts across 4 continents
https://doi.org/10.1[...]
2020
[51]
서적
Advances in the Study of Behavior
https://books.google[...]
Academic Press
2005-01-30
[52]
논문
Kleptoparasitism by great skuas (''Catharacta skua'' Brünn.) and Arctic skuas (''Stercorarius parasiticus'' L.) at a Shetland seabird colony
[53]
논문
Dimorphism, parasitism, and sex revisited: modes of reproduction among deep-sea ceratioid anglerfishes (Teleostei: Lophiiformes)
2005-08-25
[54]
서적
Online Dictionary of Invertebrate Zoology
http://slovarji.info[...]
University of Nebraska
2005
[55]
논문
Weather-mediated regulation of olive scale by two parasitoids
2001-05
[56]
웹사이트
Featured Creatures. Encarsia perplexa
http://entnemdept.uf[...]
University of Florida
2018-01-06
[57]
논문
Sex determination in Bonellia viridis (Echiura: Bonelliidae): population dynamics and evolution
https://core.ac.uk/d[...]
[58]
서적
Advances in parasitology
Elsevier Science
2011
[59]
논문
Biodiversity of hymenopteran parasitoids
2023
[60]
논문
Quantifying the unquantifiable: why Hymenoptera, not Coleoptera, is the most speciose animal order
2018-07-12
[61]
서적
Parasite Diversity and Diversification
https://books.google[...]
Cambridge University Press
[62]
서적
Modern Biology
https://books.google[...]
Pitambar Publishing
1997
[63]
논문
Developing a thief: Haustoria formation in parasitic plants
2018-10-01
[64]
서적
Parasitic flowering plants
Brill
[65]
웹사이트
Parasitic Plants of the World
http://nickrentlab.s[...]
2018-04-10
[66]
논문
Introduction to Parasitic Flowering Plants
[67]
논문
The evolution of parasitism in plants
[68]
논문
The biology of myco-heterotrophic ('saprophytic') plants
1994
[69]
논문
Biotrophic Fungal Pathogens: a Critical Overview
http://dx.doi.org/10[...]
2022-08-11
[70]
웹사이트
What is honey fungus?
https://www.rhs.org.[...]
Royal Horticultural Society
2017-10-12
[71]
논문
Biotrophy-necrotrophy switch in pathogen evoke differential response in resistant and susceptible sesame involving multiple signaling pathways at different phases
http://dx.doi.org/10[...]
2017-12-08
[72]
논문
Stop neglecting fungi
2017-07-25
[73]
논문
Epidemiology of microsporidiosis: sources and modes of transmission
https://www.research[...]
2004-12-09
[74]
논문
Transmission and epidemiology of zoonotic protozoal diseases of companion animals
2013-01
[75]
논문
Adaptive Immunity: Care for the community
2007-01
[76]
서적
Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series)
Lippincott Williams & Wilkins
[77]
논문
The ancient Virus World and evolution of cells
[78]
논문
Here a virus, there a virus, everywhere the same virus?
[79]
논문
Structural and functional studies of archaeal viruses
[80]
논문
Viral metagenomics
https://www.cbcb.umd[...]
[81]
논문
Homage to Linnaeus: How many parasites? How many hosts?
2008
[82]
논문
Common Avian Infection Plagued the Tyrant Dinosaurs
[83]
간행물
A new insect from the Cretaceous of Transbaikalia, a possible parasite of pterosaurians
1976
[84]
논문
A new flea from the Early Cretaceous of China
http://www.app.pan.p[...]
2020
[85]
논문
First discovery of parasite eggs in a vertebrate coprolite of the Late Triassic in Thailand
2023-08-09
[86]
서적
Evolution of Infectious Disease
https://archive.org/[...]
Oxford University Press
[87]
논문
The hygiene hypothesis and the increasing prevalence of chronic inflammatory disorders
https://www.academia[...]
[88]
논문
Invasion of the Gender Benders: by manipulating sex and reproduction in their hosts, many parasites improve their own odds of survival and may shape the evolution of sex itself
http://findarticles.[...]
2008-11-15
[89]
서적
What Is Life?
https://books.google[...]
Simon and Schuster
[90]
서적
A Companion to the Philosophy of Biology
https://books.google[...]
John Wiley & Sons
[91]
논문
Interference competition and parasite virulence
[92]
논문
Parasite and host assemblages: embracing the reality will improve our knowledge of parasite transmission and virulence
[93]
서적
Encyclopedia of Life Sciences
John Wiley
2006-01-27
[94]
논문
Ancient co-speciation of simian foamy viruses and primates
https://zenodo.org/r[...]
[95]
논문
Reinterpreting the origins of flamingo lice: cospeciation or host-switching?
[96]
논문
Parasite adaptation to locally common host genotypes
https://public.wsu.e[...]
[97]
논문
Altered behavior of parasitized killifish increases susceptibility to predation by bird final hosts
http://parasitology.[...]
[98]
논문
Fatal attraction in rats infected with ''Toxoplasma gondii''
[99]
논문
Evaluation of manipulative effects by an ichneumonid spider-ectoparasitoid larva upon an orb-weaving spider host (Araneidae: Cyclosa argenteoalba) by means of surgical removal and transplantation
http://dx.doi.org/10[...]
2019-09-16
[100]
서적
On the Wing: Insects, Pterosaurs, Birds, Bats and the Evolution of Animal Flight
https://books.google[...]
Oxford University Press
[101]
논문
Evolution of parasite life history traits: myths and reality
http://faculty.uml.e[...]
1995-09
[102]
논문
A cnidarian parasite of salmon (Myxozoa: Henneguya) lacks a mitochondrial genome
2020-02-19
[103]
논문
Sexual reproduction as an adaptation to resist parasites (a review).
1990-05
[104]
논문
Sex against virulence: the coevolution of parasitic diseases
[105]
논문
Parasites, Bright Males, and the Immunocompetence Handicap
https://www.academia[...]
[106]
웹사이트
Host–Parasite Interactions Innate Defenses of the Host
http://www.colorado.[...]
University of Colorado
2014-05-07
[107]
논문
Parasite immunomodulation and polymorphisms of the immune system
[108]
논문
Construction and Utilization of Multiple Combs in Polistes canadensis in Relation to the Biology of a Predaceous Moth
https://www.research[...]
[109]
논문
Plant defenses against parasitic plants show similarities to those induced by herbivores and pathogens
[110]
논문
Cross-talk between jasmonate and salicylate plant defense pathways: effects on several plant parasites
https://www.research[...]
[111]
논문
Specific and non-specific defense against parasitic attack
https://stevefrank.o[...]
[112]
논문
Plant Volatiles as a Defense against Insect Herbivores
1999-10-01
[113]
논문
Where are the parasites in food webs?
[114]
논문
Assessing the Effects of Trematode Infection on Invasive Green Crabs in Eastern North America
2015-06-01
[115]
논문
Possible Role of Fish and Frogs as Paratenic Hosts of Dracunculus medinensis, Chad
2016-08
[116]
논문
IJEE Soapbox: World free of parasites and vectors: Would it be heaven, or would it be hell?
https://people.clas.[...]
[117]
서적
The Art of being a Parasite
University of Chicago Press
[118]
논문
Is a healthy ecosystem one that is rich in parasites?
http://www.personal.[...]
[119]
논문
Everything in Moderation: Principles of Parasite Control for Wildlife Conservation
2014
[120]
논문
Parasites in food webs: the ultimate missing links
[121]
논문
Parasites in Food Webs: Untangling the Entangled Bank
[122]
논문
Quantifying parasites in samples of hosts
http://www.zoologia.[...]
[123]
논문
History of human parasitic diseases
2004-06
[124]
서적
Sept siècles de parasitologie en France
Société Française de Parasitologie
1993
[125]
논문
Francesco Redi and the birth of experimental parasitology
[126]
서적
Parasitism: The Diversity and Ecology of Animal Parasites
https://archive.org/[...]
Cambridge University Press
[127]
웹사이트
Acarus as the cause of scabies
http://pacs.unica.it[...]
University of Cagliari
2018-04-11
[128]
서적
General Parasitology
Academic Press
[129]
간행물
Parasitology
https://www.merriam-[...]
2018-04-13
[130]
논문
Sir David Bruce, a pioneer of tropical medicine
2006-03
[131]
웹사이트
Malaria and Malaria Vaccine Candidates
https://www.historyo[...]
The College of Physicians of Philadelphia
2018-02-11
[132]
뉴스
Malaria vaccine gets 'green light'
https://www.bbc.com/[...]
2015-07-24
[133]
논문
Biology and Use of the Whitefly Parasitoid Encarsia Formosa
1998
[134]
웹사이트
Parasitoid Wasps (Hymenoptera)
https://extension.um[...]
University of Maryland
2016-06-06
[135]
웹사이트
Encouraging innovation in biopesticide development.
http://ec.europa.eu/[...]
European Commission
2017-01-09
[136]
서적
24 Hours in Ancient Rome: A Day in the Life of the People Who Lived There
https://books.google[...]
Michael O'Mara
[137]
서적
The Mask of the Parasite: A Pathology of Roman Patronage
University of Michigan Press
[138]
서적
Living with Germs: In health and disease
https://books.google[...]
Oxford University Press
[139]
서적
On Poetry: A Rapsody
https://books.google[...]
And sold by J. Huggonson, next to Kent's Coffee-house, near Serjeant's-inn, in Chancery-lane; [and] at the bookseller's and pamphletshops
[140]
논문
What's in a name? Taxonomic and gender biases in the etymology of new species names
2022-05-11
[141]
서적
Networking: Communicating with Bodies and Machines in the Nineteenth Century
https://books.google[...]
University of Michigan Press
[142]
간행물
Parasitism and Symbiosis
http://www.sf-encycl[...]
2016-01-10
[143]
웹사이트
This is clearly an important species we're dealing with
http://www.deepseane[...]
Deep Sea News
2011-05-09
[144]
웹사이트
5 Alien Parasites and Their Real-World Counterparts
https://www.livescie[...]
Live Science
2012-05-29
[145]
웹사이트
Parasitism in the Alien Movies
http://www.signalton[...]
Signal to Noise Magazine
2018-02-11
[146]
뉴스
The Making of Alien's Chestburster Scene
https://www.theguard[...]
The Guardian
2010-05-29
[147]
뉴스
'Alien' Evolution: Explore Every Stage in the Xenomorph's Gruesome Life Cycle. Celebrate Alien Day with a look at the past, present and future of cinema's most terrifying extraterrestrial
http://www.indiewire[...]
IndieWire
2017-04-25
[148]
간행물
The Fungus that Reduced Humanity to The Last of Us
https://blogs.scient[...]
2013-06-25
[149]
Youtube
“寄生虫”扱い…「アンコウ」のオス、切ない生き方 『the SOCIAL』傑作選(2018年5月9日放送より)
https://www.youtube.[...]
[150]
웹사이트
第3回 寄生虫ハリガネムシはどうやって宿主の心を操るのか
https://natgeo.nikke[...]
2022-08-23
[151]
Citation
ハリガネムシは寄生したカマキリを操作し水平偏光に引き寄せて水に飛び込ませる
https://www.youtube.[...]
2021-07-09
[152]
저널
Common Avian Infection Plagued the Tyrant Dinosaurs
http://dx.doi.org/10[...]
2013-07-08
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com