하플로그룹 CT (Y-DNA)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
하플로그룹 CT (Y-DNA)는 Y 염색체의 M168 SNP를 공유하는 모든 남성을 포함하는 하플로그룹이다. 약 7~10만 년 전 아프리카에서 발생했으며, 현재 하플로그룹 A, B를 제외한 모든 현생 인류 남성이 CT의 후손이다. CT는 CF와 DE로 분화되었으며, CF는 C와 F, DE는 D와 E로 세분화된다. CT는 아프리카를 탈출한 인류의 계통을 추적하는 데 중요한 역할을 한다.

더 읽어볼만한 페이지
- 인류 Y-DNA 하플로그룹 - 하플로그룹 K (Y-DNA)
하플로그룹 K는 약 47,000~50,000년 전 중동 또는 중앙아시아에서 기원한 것으로 추정되는 Y-염색체 DNA 하플로그룹으로, 하위 계통인 L, T, NO, M, S, P 등을 통해 유라시아, 오세아니아, 아메리카 대륙 등 전 세계에 분포한다. - 인류 Y-DNA 하플로그룹 - Y염색체 아담
Y염색체 아담은 현존하는 모든 인류의 부계 혈통을 따라 올라갈 때 만나는 가장 최근의 공통 조상을 지칭하는 비유적인 용어이며, 단 한 명의 남성이 아닌 Y염색체에 돌연변이가 발생하기 이전의 남성 집단을 의미한다.
2. 발생
하플로그룹 CT에 속하는 모든 사람은 Y염색체에 P9.1, M294와 함께 M168 SNP 돌연변이 형태를 가지고 있다. M168 변이는 약 7~10만 년 전 에티오피아에서 수단에 걸친 지역의 한 남성에게서 처음 나타난 것으로 추정되며, CT의 자손이 아닌 A계통, B계통을 제외한 모든 현생인류 남성이 가지고 있다.[1][5][7]
CT 하플로그룹을 가진 남성은 Y 염색체에 SNP 돌연변이 M168을 가지고 있다. 이 돌연변이는 아프리카에서 발견되는 A와 B-M60을 제외한 거의 모든 현대 인류 남성 계통에 존재한다.[1]
CT 하플로그룹을 가진 남성은 Y 염색체에 SNP 돌연변이 M168, P9.1, M294를 가지고 있다. 이러한 돌연변이는 아프리카에서 주로 발견되는 A와 B-M60을 제외한 모든 현대 인류 남성 계통에 존재한다.[1]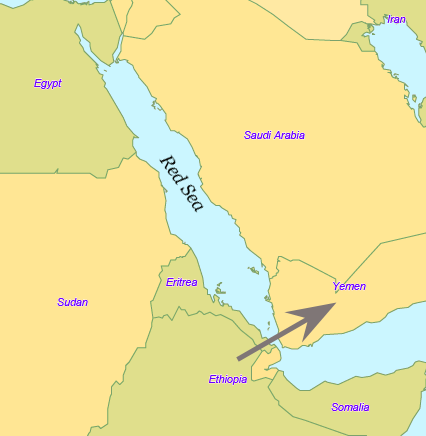
오늘날 모든 CT 남성의 가장 최근 공통 남성 계통 조상(TMRCA)은 현대 인류의 최근 아프리카 기원보다 앞섰을 것으로 추정되며, 그의 후손 중 일부는 이 이주에 참여하였다. 따라서 그는 이 이주 이전에 아프리카에 살았던 것으로 생각된다. 모든 인류의 부계 조상에게 주어진 "Y 염색체 아담"의 개념에 따라, CT-M168은 많은 아프리카 Y 계통과 함께 모든 비 아프리카 Y 계통이 그것에서 파생되었기 때문에 "유라시아 아담" 또는 "아프리카 밖 아담"으로도 불린다.[8][9][10]
현대 인구에서 파라그룹 CT* 남성은 발견된 적이 없다. 이는 이 하플로그룹을 가진 모든 남성이 여러 주요 분기 클레이드 중 하나에 속한다는 것을 의미한다. CT의 알려진 모든 생존 후손 계통은 CF와 DE 중 하나에 속한다. DE는 주로 아시아에 분포하는 D-CTS3946와 주로 아프리카에 분포하는 E-M96으로 나뉘는 반면, CF는 동아시아, 아메리카 원주민, 오세아니아의 C-M130과 대부분의 비 아프리카 인구에서 지배적인 F-M89로 나뉜다.[10]
하플로그룹 CT는 초기에 분기된 아프리카인 계통[25] 외의 모든 인류 부계의 공통 조상 남성의 Y염색체 DNA 염기 서열 SNP이며, 이 SNP를 가진 사람들(하플로그룹)의 명칭이다.
구약성서에 등장하는 인류 공통 조상 "아담"이라는 남성에 비유하여, '''"유라시안 아담"'''(유라시아 대륙에서의 Y염색체 아담)이라고 불리지만, 유라시아 대륙뿐만 아니라, 남북 아메리카 대륙과 오스트레일리아 대륙, 아프리카 대륙에 사는 흑인(E계통), 백인, 오스트레일리아 원주민, 몽골로이드의 모든 공통 조상이 되는 남성이다.
이후 하플로그룹 CT의 후손 중 DE 계통의 일부 (아프리카 동부에 남은 나머지 일부는 E 계통이 되었다) 및 CF (C 계통, I 계통, J 계통, N 계통, O 계통, R 계통, Q 계통의 조상)는 기후 변동에 따라 아프리카 대륙 동부, 현재의 지부티에서 에리트레아 부근에서 홍해 (바브엘만데브 해협)를 건너 현재의 예멘 남부에 도달하여 아프리카를 탈출했다. 이 후손들은 사하라 사막 이남 이외의 전 세계로 전파되어 번식해 나갔을 것으로 보인다.
3. 분포
오늘날 모든 CT 남성의 가장 최근 공통 남성 계통 조상(TMRCA)은 현대 인류의 최근 아프리카 기원보다 앞섰을 것이며, 그의 후손 중 일부는 이 이주에 참여했다. 따라서 그는 이 제안된 이주 이전에 아프리카에 살았던 것으로 생각된다.[5][7] CT-M168은 "Y 염색체 아담" 개념에 따라, 많은 아프리카 Y 계통과 함께 모든 비 아프리카 Y 계통이 파생되었기 때문에 "유라시아 아담" 또는 "아프리카 밖 아담" 계통으로도 언급된다.[8][9][10]
현대 인구에서 파라그룹 CT* 남성은 발견된 적이 없다. 이 하플로그룹을 가진 모든 남성은 여러 주요 분기 클레이드 중 하나에 속한다. CT의 알려진 모든 생존 후손 계통은 CF와 DE 두 개의 주요 하위 클레이드 중 하나에 속한다. DE는 주로 아시아에 분포하는 하플로그룹 D-CTS3946와 주로 아프리카에 분포하는 하플로그룹 E-M96으로 나뉘며, CF는 동아시아, 아메리카 원주민, 오세아니아의 하플로그룹 C-M130과 대부분의 비 아프리카 인구에서 지배적인 하플로그룹 F-M89로 나뉜다.[10]
하플로그룹 CT를 정의하는 변이 "M168"은 약 6만 년에서 7만 9천 년 전 쯤에 에티오피아에서 수단 부근에 살았던 한 남성에게서 발생했다. Y 염색체 상의 DNA염기 서열 변이 "M168"은 일부 아프리카인을 제외한 모든 인류의 남성이 물려받았다. 이는 아프리카인의 일부 하플로그룹 "A 계통"과 "B 계통"이 그 이전에 분기된 종족임을 나타낸다.
이후 하플로그룹 CT의 후손 중 DE 계통의 일부 (아프리카 동부에 남은 나머지 일부는 E 계통이 되었다) 및 CF (C 계통이나 I 계통, J 계통, N 계통, O 계통, R 계통, Q 계통의 조상)는 기후 변동에 따라 아프리카 대륙 동부, 현재의 지부티에서 에리트레아 부근에서 홍해 (바브엘만데브 해협)를 건너 현재의 예멘 남부에 도달하여 아프리카를 탈출했다. 이 후손들은 사하라 사막 이남 이외의 전 세계로 퍼져나가 번식했을 것으로 보인다.
4. 하위 계통
오늘날 모든 CT 남성의 가장 최근 공통 남성 계통 조상(TMRCA)은 현대 인류의 최근 아프리카 기원보다 앞섰을 것으로 추정되며, 그의 후손 중 일부는 이 이주에 참여했을 것으로 보인다. 따라서 그는 이 이주 이전에 아프리카에 살았던 것으로 생각된다.[1][5][7] CT-M168은 모든 비 아프리카 Y 계통이 파생된 기원이기 때문에, "Y 염색체 아담" 개념과 관련하여 "유라시아 아담" 또는 "아프리카 밖 아담" 계통으로 언급되기도 한다.[8][9][10]
현대 인구에서 파라그룹 CT* 남성은 발견된 적이 없다. 즉, 이 하플로그룹을 가진 모든 남성은 CF 또는 DE 중 하나에 속한다.
4. 1. 하플로그룹 CF (Y-DNA)
하플로그룹 CF는 하플로그룹 CT의 하위 그룹이다.
하플로그룹 CT를 정의하는 변이 "M168"은 약 6만 년에서 7만 9천 년 전 쯤에 에티오피아에서 수단 부근에 살았던 한 남성에게서 발생했다. Y 염색체 상의 DNA염기 서열 변이 "M168"을 일부 아프리카인을 제외한 모든 인류의 남성이 물려받았다. 이는 아프리카인의 일부 하플로그룹 "A 계통"과 "B 계통"이 그 이전에 분기된 종족임을 나타낸다.
이후 하플로그룹 CT의 후손 중 DE 계통의 일부 (아프리카 동부에 남은 나머지 일부는 E 계통이 되었다) 및 CF (C 계통이나 I 계통, J 계통, N 계통, O 계통, R 계통, Q 계통의 조상)는 기후 변동에 따라 아프리카 대륙 동부, 현재의 지부티에서 에리트레아 부근에서 홍해 (바브엘만데브 해협)를 건너 현재의 예멘 남부에 도달하여 아프리카를 탈출했다. 이 후손들은 사하라 사막 이남 이외의 전 세계로 전파되어 번식해 나갔을 것으로 보인다.
하플로그룹 CF의 하위 그룹은 다음과 같다.4. 2. 하플로그룹 DE
하플로그룹 DE는 하플로그룹 CT의 하위 분류이다.
하플로그룹 CT를 정의하는 변이 "M168"은 약 6만 년에서 7만 9천 년 전 쯤에 에티오피아에서 수단 부근에 살았던 한 남성에게서 발생했다. Y 염색체 상의 DNA염기 서열 변이 "M168"을 일부 아프리카인을 제외한 모든 인류의 남성이 물려받았다. 이는 아프리카인의 일부 하플로그룹 "A 계통"과 "B 계통"이 그 이전에 분기된 종족임을 나타낸다.
이후 하플로그룹 CT의 후손 중 DE 계통의 일부 (아프리카 동부에 남은 나머지 일부는 E 계통이 되었다) 및 CF (C 계통, I 계통, J 계통, N 계통, O 계통, R 계통, Q 계통의 조상)는 기후 변동에 따라 아프리카 대륙 동부, 현재의 지부티에서 에리트레아 부근에서 홍해 (바브엘만데브 해협)를 건너 현재의 예멘 남부에 도달하여 아프리카를 탈출했다. 이 후손들은 사하라 사막 이남 이외의 전 세계로 전파되어 번식해 나갔을 것으로 보인다.
하플로그룹 DE는 나이지리아, 기니비사우, 카리브 제도, 티베트 등지에서 발견된다.
5. 계통도
이 하플로그룹 집단에 속하는 모든 사람은 Y염색체에 P9.1, M294와 함께 M168 SNP 돌연변이 형태를 가지고 있다. M168 변이는 현대로부터 약 7~10만 년 전 정도에 현재의 에티오피아에서 수단에 걸친 지역의 한 남성에게서 처음 나타난 것으로 추정되며, CT의 자손이 아닌 A 계통, B 계통에 속하는 자를 제외한 모든 현생인류 남성이 가지고 있다.[1]
CT 하플로그룹을 가진 남성은 P9.1 및 M294와 함께 Y 염색체에 SNP 돌연변이 M168을 가지고 있다. 이러한 돌연변이는 거의 대부분 아프리카에서 발견되는 A와 B-M60을 제외한 모든 현대 인류 남성 계통에 존재한다.
오늘날 모든 CT 남성의 가장 최근 공통 남성 계통 조상(TMRCA)은 아마도 현대 인류의 최근 아프리카 기원보다 앞섰을 것이며, 그의 후손 중 일부는 이 이주에 참여했다. 따라서 그는 이 제안된 이주 이전에 아프리카에 살았던 것으로 생각된다.[1][5][7] 살아있는 모든 인류의 부계 조상에게 주어진 "Y 염색체 아담"의 개념에 따라, CT-M168은 많은 아프리카 Y 계통과 함께 모든 비 아프리카 Y 계통이 그것에서 파생되었기 때문에 대중화된 설명에서 "유라시아 아담" 또는 "아프리카 밖 아담"의 계통으로도 언급되었다.[8][9][10]
현대 인구에서 파라그룹 CT* 남성은 발견된 적이 없다. 이는 이 하플로그룹을 가진 모든 남성이 여러 주요 분기 클레이드 중 하나에 속한다는 것을 의미한다. CT의 알려진 모든 생존 후손 계통은 두 개의 주요 하위 클레이드인 CF와 DE 중 하나에 속한다. 차례로 DE는 주로 아시아에 분포하는 D-CTS3946와 주로 아프리카에 분포하는 E-M96으로 나뉘는 반면, CF는 동아시아, 아메리카 원주민, 오세아니아 C-M130과 대부분의 비 아프리카 인구에서 지배적인 F-M89로 나뉜다.[10]
;하플로그룹 CT (M168/PF1416)
- CF (P143/PF2587, CTS3818/M3690/PF2668, CTS6376/M3711/PF2697) 샨족
- *C (M130/Page51/RPS4Y711, M216) '' 아시아, 오세아니아, 그리고 북아메리카''
- ** C1 F3393/Z1426
'''C1a''' CTS11043
* '''C1a1''' ''이전 명칭 C1'' (M8, M105, M131) '' 일본''
* '''C1a2''' ''서유라시아에서 매우 낮은 빈도 (38,000년 전 코스텐키)''
'''C1b'''
* '''C1b1'''
**'''C1b1a'''
***'''C1b1a1''' ''이전 명칭 C5'' (M356) '' 남아시아, 중앙아시아, 그리고 서남아시아''
* '''C1b2'''
**'''C1b2a''' ''이전 명칭 C2'' (M38) '' 인도네시아, 뉴기니, 멜라네시아, 미크로네시아, 그리고 폴리네시아''
**'''C1b2b''' ''이전 명칭 C4'' (M347) '' 호주 원주민''
- **C2 ''이전 명칭 C3'' (M217, P44) ''유라시아와 북아메리카 전역에서 발견되지만, 특히 몽골인, 카자흐인, 퉁구스족, 고대 시베리아인, 그리고 나데네어를 사용하는 민족에게서 두드러짐''
- *F (M89, M213) ''유라시아, 오세아니아, 그리고 아메리카 전역에서 발견됨''
- **'''F1''' (P91, P104)
- **'''F2''' (M427, M428)
- **'''F3''' (P96)
- **'''F4''' (M481)
- *GHIJK F1329/M3658/PF2622/YSC0001299, CTS2254/M3680/PF2657, FGC2045/Z12203 유라시아 전역, 또한 아메리카 대륙 ; 아프리카와 오세아니아에서는 낮은 수준/소수 민족에게서 발견
- **G M201, P257 ''주로 코카서스; 또한 유럽, 북아프리카, 서남아시아, 중앙아시아에서 낮은 수준/소수 민족에게서 발견''
- **HIJK 유라시아 전역, 또한 아메리카 대륙; 아프리카와 오세아니아에서는 낮은 수준/소수 민족에게서 발견
H M69, M370 '' 남아시아, 중앙아시아, 그리고 서남아시아''
IJK L15/S137, L16/S138 '' 유라시아, 북아프리카, 오세아니아, 아메리카 대륙, 동아프리카''
IJ M429/P125 '' 유럽, 서아시아, 북아프리카 및 동아프리카''
*I M170, M258, P19, P38, P212, U179 '' 유럽''
*J 12f2.1, M304 '' 유럽, 서아시아, 남아시아, 북아프리카 및 동아프리카''
K M9 ''유라시아 전역에서 발견; 또한 아메리카 대륙과 오세아니아에서 유의미하며, 북아프리카와 동아프리카에서는 낮은 수준으로 존재''
*LT ('''K1''') L298/P326
**L M11, M20, M22, M61, M185, M295 '' 인도 아대륙''
**T M70, M184/USP9Y+3178, M193, M272 (이전 명칭 K2) '' 서남아시아, 남아시아, 북아프리카, 아프리카의 뿔, 차드 분지, 그리고 남유럽''
*K2 M526 유라시아, 오세아니아, 아메리카 대륙, 일부 호주 원주민과 아프리카의 일부 소수 민족
**NO ('''K2a''') M214 ''부이족과 일본 남성에게서 소수 발견되었다고 보고됨.''
***N M231 '' 동유럽, 북아시아, 그리고 동아시아''
***O M175 '' 오세아니아, 동남아시아, 그리고 동아시아''
**K2b ''이전 명칭 MPS'' P331 ''유라시아, 아메리카 대륙, 오세아니아, 그리고 아프리카의 소수 민족''
***K2b1 ''이전 명칭 MS'' P397/P399 멜라네시아인, 미크로네시아인, 호주 원주민 및 폴리네시아인.
M P256 ''인도네시아 동부, 파푸아뉴기니, 그리고 멜라네시아''
S M230, P202, P204 (이전 명칭 K5) ''인도네시아 동부, 파푸아뉴기니, 그리고 멜라네시아''
***P ('''K2b2''') 92R7, M45, M74/N12, P27.1/P207 ''유라시아, 아메리카 대륙 전역, 그리고 아프리카의 일부 소수 민족''
Q M242 '' 아메리카 대륙과 유라시아''
R M207/UTY2, M306/S1 ''유럽, 근동, 남아시아, 차드 분지, 카나리아 제도''
**K2c P261 ''발리에서 낮은 수준''
**K2d P402/P403 ''자바에서 낮은 수준''
**K2e M147 ''남아시아에서 낮은 수준''
- 하플로그룹 DE (M1/YAP, M145/P205, M203/Page36, P144, P153, P165, P167, P183) ''아시아, 아프리카, 남유럽 및 동유럽; 또한 오세아니아에서 낮은 수준''
- *하플로그룹 D (M174) ''주로 일본, 티베트 및 안다만 제도 거주자; 또한 동아시아, 중앙아시아, 미크로네시아, 멜라네시아에서 낮은 수준/소수 민족에게서 발견''
- **하플로그룹 D1 (CTS11577) ''주로 티베트; 중앙아시아, 동아시아, 동남아시아의 소수 민족''
하플로그룹 D1a Z27276 ''동아시아 및 중앙아시아''
*D1a1 (M15) (구 D1) ''강족; 또한 동아시아 및 중앙아시아 전역에서 낮은 수준''
* 하플로그룹 D1a2 P99 (구 D3) ''티베트 및 중앙아시아''
하플로그룹 D1b (구 D2) (M64.1/Page44.1, M55, M57, M179/Page31, M359.1/P41.1, P37.1, P190, 12f2.2) ''주로 일본; 또한 한국, 중국, 미크로네시아 및 멜라네시아에서 낮은 수준/소수 민족에게서 발견''
- **하플로그룹 D2 L1366 (구 D1a) ''필리핀''
- *하플로그룹 E (M40, M96) ''아프리카, 중동, 남유럽 및 동유럽''
- **하플로그룹 E1 (P147) ''아프리카 전역에서 높은 수준; 중동 및 유럽에서 낮은 수준''
하플로그룹 E1a (이전 명칭 E1) (M132) ''주로 아프리카''
하플로그룹 E1b (P177/PF1939)
*하플로그룹 E1b1 (이전 명칭 E3) (P2, DYS391p)
**하플로그룹 E1b1a (V38)
**하플로그룹 E1b1a1 (이전 명칭 E3a) (M2) '' 서아프리카, 중앙아프리카, 동남아프리카 및 남아프리카''
**하플로그룹 E1b1b (이전 명칭 E3b) (M215) '' 에티오피아인, 소말리아인, 에리트레아를 포함한 아프리카의 뿔, 북아프리카 (베르베르인, 아랍인[11]), 중동, 유럽 (특히 지중해 인근 지역)''
- **하플로그룹 E2 (M75) ''동아프리카''
6. 관련 유전학
인간 Y 염색체 DNA 하플로그룹, 세계 인구의 Y 염색체 하플로그룹, 민족별 Y-DNA 하플로그룹, 동아시아 및 동남아시아 인구의 Y-DNA 하플로그룹, 오세아니아 인구의 Y-DNA 하플로그룹과 관련된 하플로그룹 CT의 하위 그룹은 다음과 같다.
참조
[1]
논문
New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree
http://www.genome.or[...]
[2]
논문
A recent bottleneck of Y chromosome diversity coincides with a global change in culture
2015-04
[3]
논문
A Rare Deep-Rooting D0 African Y-chromosomal Haplogroup and its Implications for the Expansion of Modern Humans Out of Africa
2019-06
[4]
논문
Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations
[5]
서적
Genes, Culture, and Human Evolution
Wiley
[6]
논문
A recent bottleneck of Y chromosome diversity coincides with a global change in culture
2015-04
[7]
논문
Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations
[8]
서적
Genes, Culture, and Human Evolution: A Synthesis
https://books.google[...]
[9]
서적
Darwinian Detectives: Revealing the Natural History of Genes and Genomes
https://books.google[...]
[10]
논문
New Binary Polymorphisms Reshape and Increase Resolution of the Human Y-Chromosomal Haplogroup Tree
http://www.genome.or[...]
[11]
논문
Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel
http://www.nature.co[...]
2010
[12]
논문
Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome
[13]
웹사이트
Y-DNA Haplogroup Tree 2015
http://www.isogg.org[...]
2015-02-01
[14]
문서
Haplogroup A0-T is also known as A-L1085 (and previously as A0'1'2'3'4).
[15]
문서
Haplogroup A1 is also known as A1'2'3'4.
[16]
문서
F-Y27277, sometimes known as F2'4, is both the parent clade of F2 and F4 and a child of F-M89.
[17]
문서
Haplogroup LT (L298/P326) is also known as Haplogroup K1.
[18]
문서
Between 2002 and 2008, [[Haplogroup T-M184]] was known as "Haplogroup K2". That name has since been re-assigned to [[Haplogroup K2|K-M526]], the sibling of Haplogroup LT.
[19]
문서
Haplogroup K2b (M1221/P331/PF5911) is also known as Haplogroup MPS.
[20]
문서
Haplogroup K2b1 (P397/P399) is also known as Haplogroup MS, but has a broader and more complex internal structure.
[21]
문서
Haplogroup P (P295) is also klnown as K2b2.
[22]
웹사이트
K-M2335
https://www.yfull.co[...]
2017
[23]
문서
Haplogroup S, as of 2017, is also known as K2b1a. (Previously the name Haplogroup S was assigned to K2b1a4.)
[24]
문서
Haplogroup M, as of 2017, is also known as K2b1b. (Previously the name Haplogroup M was assigned to K2b1d.)
[25]
문서
ハプログループは「A」と「B」はCTの子孫ではない。CTの子孫のうち、「E」以外の系統は出アフリカを果たし、「E」はアフリカにとどまった。すなわち「A」、「B」、「E」が非出アフリカ組、「D」、「CF」が出アフリカ組である。
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com