프로테아좀
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
프로테아좀은 단백질 분해 효소 활성을 가진 거대 복합체로, 세포 내 단백질 항상성 유지에 핵심적인 역할을 한다. 26S 프로테아좀은 20S 코어 입자와 양쪽에 결합된 19S 조절 입자로 구성되며, 11S 조절 입자(PA28) 또는 PA200/Blm10이 20S에 결합하기도 한다. 프로테아좀은 유비퀴틴화된 단백질을 인식하여 분해하며, 세포 주기 조절, 세포 신호 전달, 면역 반응 등 다양한 생물학적 과정에 관여한다. 프로테아좀 기능 이상은 신경퇴행성 질환, 심혈관 질환, 암, 자가면역 질환 등 다양한 질병과 관련이 있으며, 프로테아좀 억제제는 항암 치료 등에 사용된다.

더 읽어볼만한 페이지
- 단백질 복합체 - 단백질-단백질 상호작용
단백질-단백질 상호작용은 단백질 간의 결합을 의미하며, 결합의 지속성, 결합력, 구성 단백질 종류에 따라 다양한 유형으로 분류되고, 물 분자에 의해 조절되며, 연구 방법과 데이터베이스를 통해 질병 연구 및 신약 개발에 응용된다. - 단백질 복합체 - 광계 I
광계 I(PSI)은 광합성 전자전달계의 효소로, 빛 에너지를 흡수하여 전자를 전달하며, P700을 포함한 다양한 색소, 조효소, 보조 인자로 구성되어 있다. - 생물학 - 균근
균근은 식물 뿌리와 균류 사이의 공생체로, 식물에게 물과 영양분을 공급하고 식물은 광합성 산물을 제공하며, 농업, 산림 복원 등 다양한 분야에서 활용될 수 있다. - 생물학 - 생명
생명은 환경 속에서 자기 유지, 증진, 강화를 하는 특징으로 정의되나 논쟁적이며, 항상성 유지, 세포 구조, 물질대사, 성장, 적응, 자극 반응, 생식 능력을 포함하고, 자기 복제와 진화로 유지되는 열린 열역학계로 설명되며, 기원은 약 40억 년 전으로 추정되고, 진화와 멸종이 공존하며, 바이러스, 외계 생명체, 인공 생명 연구가 진행 중이다.

2. 구조

26S 프로테아좀은 프로테아제 활성을 가진 관통형의 20S 프로테아좀 양쪽에 19S 복합체가 2개 결합한 구조이다. 20S 프로테아좀, 19S 복합체, 11S 복합체에 대해 자세히 알아보자.
- 20S 프로테아좀 (CP : core particle)
: 20S 프로테아좀은 α 소단위 7분자로 구성된 α-ring과 β 소단위 7분자로 구성된 β-ring이 αββα 순으로 겹쳐진 관통 구조를 하고 있다. 안의 빈 공간은 단백질 분해 장소로 β1, β2, β5가 각각 트립신(trypsin영어), 키모트립신(chymotrypsin영어), 펩타이딜-글루타밀 펩타이드(peptidyl-glutamyl peptide영어) 가수분해(PGPH) 활성을 가지고 있다. 보통 20S 프로테아좀의 상태로는 α-ring이 닫혀 있어 기질이 안에 들어갈 수 없다. 20S 프로테아좀 분자집합은 PAC1, 2, 3, 4(Pba1, 2, 3, 4), Ump1 등 분자 샤페론에 의해 정상적으로 조립된다.[144]
- 19S 복합체 (RP : regulatory particle, PA700)
: 관통형 20S 프로테아좀 양끝에 결합해 뚜껑 역할을 한다. 19S 복합체는 base와 lid로 구성되어 있다. Base는 Rpt1-6와 Rpn1-2의 8개 단백질 분자로 구성되어 20S 프로테아좀의 α-ring 개폐를 제어하고, 표적단백질을 펴는 것(unfolding영어)에 관여한다. lid는 Rpn3, 5-9, 11, 12의 8개 단백질 분자로 구성되어 탈유비퀴틴화 반응에 관여한다. 또, base와 lid를 연결하는 Rpn10 소단위는 표적단백질의 인식, 포착에 관여한다.
- 11S 복합체 (PA28)
: 11S 복합체는 19S 복합체와 같이 20S 프로테아좀의 양 끝에 결합하는 8량체이다. ATP가수분해효소 활성을 가진 소단위를 가지고 있지 않고, 짧은 펩타이드 분해에 기여한다.
26S 프로테아좀은 프로테아제 활성을 갖는 원통형의 20S 프로테아좀에 그 덮개 역할을 하는 19S 복합체가 2개 결합한 것으로, "덤벨형"이라고도 한다. 그에 비해 20S 프로테아좀의 양쪽에 11S 복합체가 각각 결합한 것은 "풋볼형"이라고 불린다.
2. 1. 20S 핵심 입자 (CP)
20S 코어 입자(Core Particle, CP)는 프로테아좀의 핵심 부분으로, 단백질 분해 기능을 담당한다.
20S 코어 입자는 α 소단위체 7개로 구성된 α-ring과 β 소단위체 7개로 구성된 β-ring이 αββα 순서로 겹쳐진 원통형 구조를 하고 있다.
- 모든 20S 입자는 두 가지 유형의 소단위체로 구성된 4개의 적층된 7량체 고리 구조로 구성된다.
- α 소단위체는 본질적으로 구조적이며, β 소단위체는 주로 촉매 작용을 한다.
- α 소단위체는 β 소단위체와 상동성을 갖는 유사 효소이다.
- 바깥쪽 두 고리는 각각 7개의 α 소단위체로 구성되어 있으며, 조절 입자 및 α 소단위체의 N-말단에 대한 도킹 도메인 역할을 하여, 기질이 내부 공동으로 접근하는 것을 차단하는 게이트를 형성한다.[17]
- 안쪽 두 고리는 각각 7개의 β 소단위체로 구성되어 있으며, N-말단에는 단백질 분해 반응을 수행하는 단백질 분해 효소 활성 부위가 포함되어 있다.[18]
안쪽의 빈 공간은 단백질 분해 장소이며, β1, β2, β5 소단위체가 각각 트립신(trypsin영어), 키모트립신(chymotrypsin영어), 펩타이딜-글루타밀 펩타이드(peptidyl-glutamyl peptide영어) 가수분해(PGPH) 활성을 가지고 있어, 각기 다른 촉매 활성을 담당한다.[140]
일반적으로 20S 프로테아좀만으로는 α-ring이 닫혀 있어 기질(분해될 단백질)이 안으로 들어갈 수 없다. 20S 프로테아좀 분자 집합체는 PAC1, 2, 3, 4(Pba1, 2, 3, 4), Ump1 등의 분자 샤페론에 의해 정상적으로 조립된다.[144]
고세균의 ''Thermoplasma acidophilum''과 같은 경우, 모든 α 및 β 소단위체가 동일하며, 효모와 같은 진핵생물 프로테아좀은 각 소단위체에 대해 7가지 뚜렷한 유형을 포함한다. 포유류에서 β1, β2, β5 소단위체는 촉매 작용을 한다. 비록 공통적인 메커니즘을 공유하지만, 키모트립신 유사, 트립신 유사 및 펩티딜-글루타밀 펩타이드-가수분해 (PHGH)로 간주되는 세 가지 뚜렷한 기질 특이성을 갖는다.[21]
프로테아좀의 크기는 비교적 보존되어 있으며 약 150 옹스트롬(Å) x 115 Å이다. 내부 공간은 최대 53 Å 너비이지만, 입구는 13 Å만큼 좁을 수 있으며, 이는 기질 단백질이 들어가려면 최소한 부분적으로 펼쳐져야 함을 시사한다.[20]
β 소단위체는 N-말단 "프로펩티드"와 함께 합성되어 활성 부위 노출을 위해 번역 후 변형을 거친다. 20S 입자는 두 개의 반쪽 프로테아좀(7개 α-ring + 7개 프로-β 링)으로 조립된다. 두 반쪽의 β 링 결합은 프로펩티드의 트레오닌 의존적 자가분해를 유발, 활성 부위를 노출시킨다. 이러한 β 상호작용은 주로 염다리와 알파 나선 간 소수성 상호작용으로 매개되며, 돌연변이에 의해 파괴되면 조립이 손상된다.[43]
최근에는 α3 코어 소단위체가 없는 인간 세포에서 대체 프로테아좀이 확인되었다.[22] 이러한 프로테아좀(α4-α4 프로테아좀)은 누락된 α3 소단위체 대신 추가 α4 소단위체를 포함하는 20S 코어 입자를 형성한다. 이러한 대체 'α4-α4' 프로테아좀은 이전에 효모에서 존재하는 것으로 알려져 있었다.[23] 이러한 프로테아좀 이소형의 정확한 기능은 여전히 대부분 알려지지 않았지만, 이러한 프로테아좀을 발현하는 세포는 카드뮴과 같은 금속 이온에 의해 유도된 독성에 대한 내성이 향상된다는 것을 보여준다.[22][24]
2. 2. 19S 조절 입자 (RP)
19S 조절 입자(regulatory particle, RP, PA700)는 원통형 20S 프로테아좀 양 끝에 결합하여 뚜껑 역할을 한다. 19S 복합체는 base(기저부)와 lid(뚜껑부)로 구성되어 있다. Base는 Rpt1-6와 Rpn1-2의 8개 단백질 분자로 구성되어 20S 프로테아좀의 α-ring 개폐를 제어하고, 표적단백질을 펴는 것(unfolding영어)에 관여한다. lid는 Rpn3, 5-9, 11, 12의 8개 단백질 분자로 구성되어 탈유비퀴틴화 반응에 관여한다. 또, base와 lid를 연결하는 Rpn10 소단위는 표적단백질의 인식, 포착에 관여한다.[144]진핵생물의 19S 입자는 19개의 개별 단백질로 구성되어 있으며, 20S 코어 입자의 α 링에 직접 결합하는 9개의 서브유닛 염기와 10개의 서브유닛 뚜껑의 두 개의 하위 조립체로 나눌 수 있다. 9개의 염기 단백질 중 6개는 AAA 계열의 ATPase 서브유닛이며, 이러한 ATPase의 진화적 동족체는 고세균에 존재하며, PAN(프로테아좀 활성화 뉴클레오티데이스)이라고 한다.[25] 19S와 20S 입자의 결합에는 ATP가 19S ATPase 서브유닛에 결합하는 것이 필요하며, ATP 가수 분해는 조립된 복합체가 접힌 및 유비퀴틴화된 단백질을 분해하는 데 필요하다. 기질의 언폴딩 단계만이 ATP 가수 분해로부터 에너지를 필요로 하며, ATP 결합만으로는 단백질 분해에 필요한 다른 모든 단계(예: 복합체 조립, 게이트 열림, 전위 및 단백질 분해)를 지원할 수 있다.[26][27] 실제로, ATP가 ATPases에 결합하는 것 자체만으로도 언폴딩된 단백질의 빠른 분해를 지원한다. 그러나 ATP 가수 분해가 언폴딩에만 필요하지만, 이 에너지가 이러한 단계 중 일부의 결합에 사용될 수 있는지는 아직 명확하지 않다.[27][28]
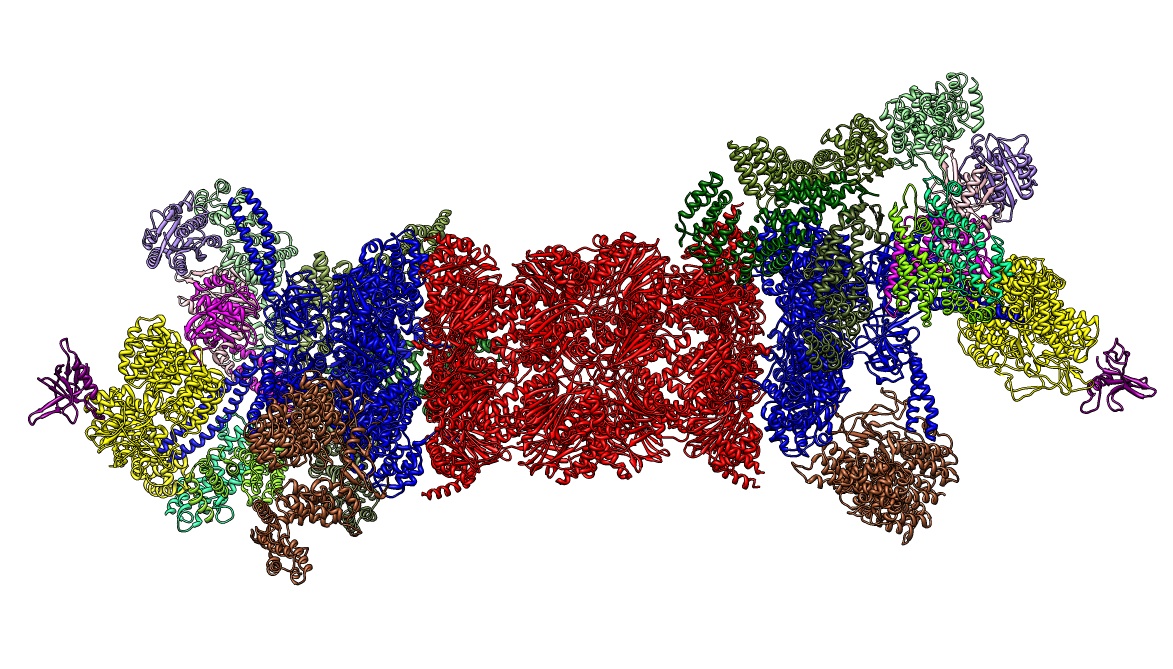
2012년, 단일 입자 분석을 통해 26S 프로테아좀의 분자 구조가 밝혀졌다.[30][31] 2016년에는 크라이오-EM을 통해 기질이 없는 상태에서 인간 26S 프로테아좀의 첫 번째 원자 수준 구조가 결정되었다.[32][33][34] 2018년에는 기질이 결합된 26S 프로테아좀의 7개의 원자 구조를 동시에 결정하여 탈유비퀴틴화, 전위 시작 및 기질의 순차적 언폴딩의 상세한 메커니즘을 밝히는 주요 노력이 이루어졌다.[15] 19S의 중심부에는 20S에 직접 인접하여 Rpt1/Rpt2/Rpt6/Rpt3/Rpt4/Rpt5 순서의 이종 육각형 링으로 조립되는 AAA-ATPases (AAA 단백질)가 있다. 이 링은 이량체의 삼량체이다: Rpt1/Rpt2, Rpt6/Rpt3 및 Rpt4/Rpt5는 N-말단 코일 코일을 통해 이량체를 형성한다. 이러한 코일 코일은 육각형 링에서 돌출되어 있다. 가장 큰 조절 입자 비-ATPases Rpn1 및 Rpn2는 각각 Rpt1/2 및 Rpt6/3의 끝에 결합한다. 유비퀴틴 수용체 Rpn13은 Rpn2에 결합하여 염기 서브 복합체를 완성한다. 뚜껑은 AAA-ATPase 육합체(Rpt6/Rpt3/Rpt4)의 절반을 덮고 있으며, Rpn6을 통해, 그리고 더 적은 정도는 Rpn5를 통해 20S와 직접 접촉한다. 구조적으로 상호 관련이 있으며 COP9 신호소 및 eIF3의 서브유닛(따라서 PCI 서브유닛이라고 함)인 서브유닛 Rpn9, Rpn5, Rpn6, Rpn7, Rpn3 및 Rpn12는 Rpn8/Rpn11 이량체를 둘러싸는 말굽 모양의 구조로 조립된다. 탈유비퀴틴화 효소인 Rpn11은 AAA-ATPase 육합체의 입구에 위치하여 기질이 20S로 전위되기 직전에 유비퀴틴 잔기를 제거하는 데 이상적으로 위치한다. 현재까지 확인된 두 번째 유비퀴틴 수용체인 Rpn10은 뚜껑의 주변, 서브유닛 Rpn8 및 Rpn9 근처에 위치한다.
26S 프로테아좀 전체 효소 내의 19S 조절 입자는 현재까지 기질이 없는 상태에서 6가지의 매우 다른 형태적 상태로 관찰되었다.[35][36] 이 지배적인 저에너지 상태에서 AAA-ATPase 구성의 특징은 AAA-도메인의 계단 또는 록와셔와 같은 배열이다.[29][30] ATP가 존재하지만 기질이 없는 상태에서는 19S의 3가지 다른, 덜 흔한 형태가 채택되며, 이는 주로 AAA-ATPase 모듈에 대한 뚜껑의 위치에 따라 다르다.[32][36] ATP-γS 또는 기질이 존재할 때 AAA-ATPase 모듈의 극적인 구조적 변화를 나타내는 훨씬 더 많은 형태가 관찰되었다.[15][35][37][38] 일부 기질 결합 형태는 기질이 없는 형태와 매우 유사하지만, 특히 AAA-ATPase 모듈에서 완전히 동일하지는 않다.[15][35] 26S 조립 전, 자유 형태의 19S 조절 입자는 7가지 형태적 상태에서도 관찰되었다.[39] 특히, 이러한 모든 컨포머는 다소 다르며 뚜렷한 특징을 나타낸다. 따라서 19S 조절 입자는 다양한 생리적 조건에서 최소 20개의 형태적 상태를 샘플링할 수 있다.
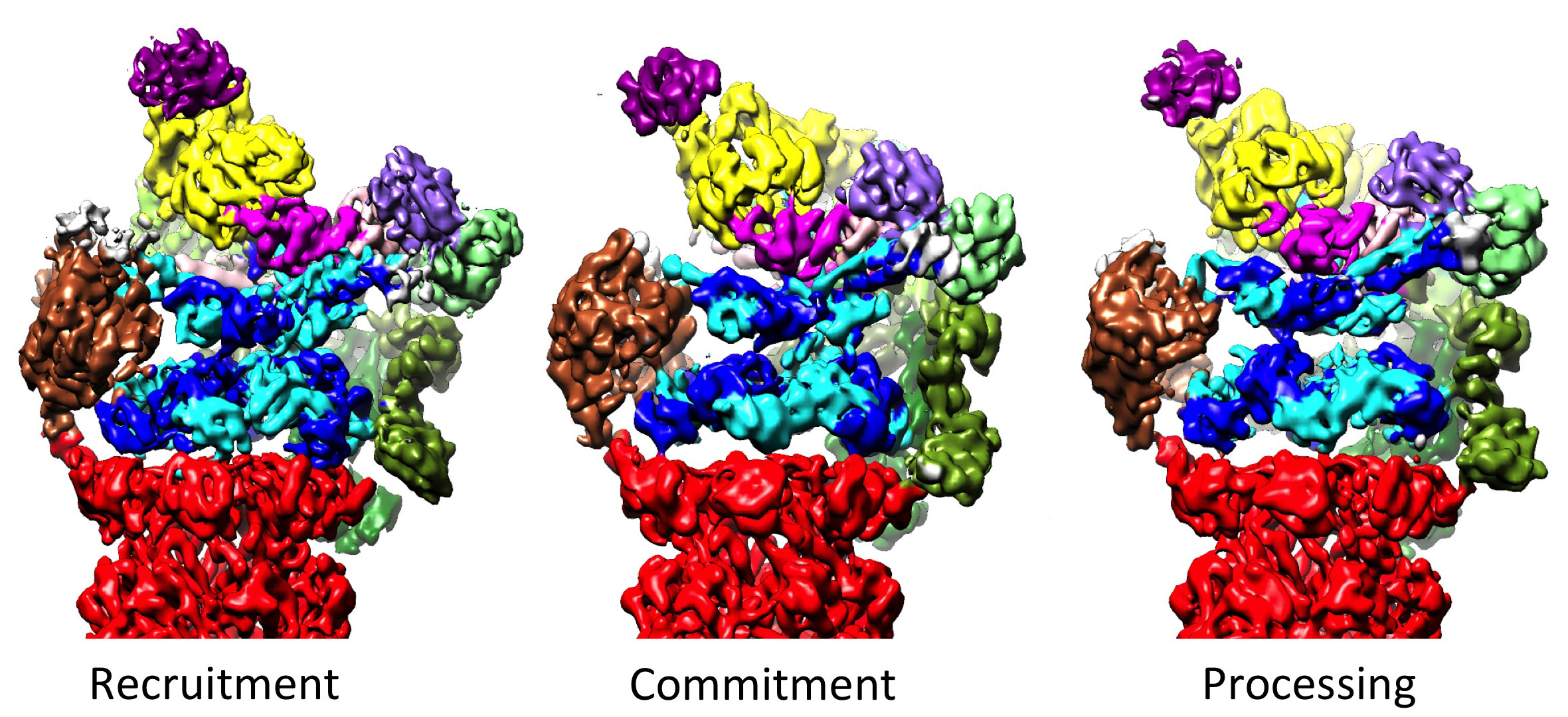
19S 조절 입자는 20S를 자극하여 단백질을 분해하는 역할을 한다. 19S 조절 ATPases의 주요 기능은 기질이 분해 챔버로 들어가는 것을 막는 20S의 게이트를 여는 것이다.[40] 프로테아좀 ATPase가 이 게이트를 여는 메커니즘은 최근에 밝혀졌다.[17] 20S 게이트 개방에는 특정 모티프 (예: HbYX 모티프)를 포함하는 프로테아좀 ATPases의 C-말단이 필요하다. ATPases C-말단은 20S 상단의 포켓에 결합하여 ATPase 복합체를 20S 단백질 분해 복합체에 묶어 기질 펼침 장치를 20S 분해 기계에 연결한다. 이러한 C-말단이 20S 포켓에 결합하는 것만으로도 20S의 게이트가 열리는 것이 자극된다.[17] 이러한 "열쇠-잠금" 메커니즘이 작동하는 정확한 메커니즘은 인간 26S 프로테아좀의 맥락에서 원자 수준에 가까운 해상도로 구조적으로 규명되었으며, ATPase 서브유닛 Rpt1/2/3/5/6의 5개의 C-말단이 20S 표면 포켓에 삽입되어야 20S 게이트가 완전히 열리는 것으로 나타났다.[35][15][32]
19S 조절 입자의 조립 과정은 최근에야 상당히 해명되었다. 19S 조절 입자는 베이스와 뚜껑이라는 두 개의 별개의 서브 구성 요소로 조립된다. 베이스 복합체의 조립은 4개의 조립 샤페론인 Hsm3/S5b, Nas2/p27, Rpn14/PAAF1, Nas6/간키린(효모/포유류의 이름)에 의해 촉진된다.[45] 이 조립 샤페론은 AAA-ATPase 서브유닛에 결합하며, 이들의 주요 기능은 이종 6량체 AAA-ATPase 링의 적절한 조립을 보장하는 것으로 보인다. 현재까지 베이스 복합체가 별도로 조립되는지, 20S 코어 입자에 의해 조립이 템플릿화되는지, 아니면 대체 조립 경로가 존재하는지에 대한 논쟁이 여전히 진행 중이다. 4개의 조립 샤페론 외에도 탈유비퀴틴화 효소 Ubp6/Usp14 또한 베이스 조립을 촉진하지만 필수적이지는 않다.[46] 뚜껑은 특정 순서로 별도로 조립되며 조립 샤페론이 필요하지 않다.[47]
2. 3. 11S 조절 입자 (PA28)
11S복합체(PA28)는 19S복합체와 마찬가지로 20S프로테아좀의 양 끝에 결합하는 8량체이다. ATPase활성을 가진 소단위를 가지고 있지 않고, 짧은 펩타이드 분해에 기여한다.[41] 11S 입자는 바이러스 감염 후 생성된 것과 같은 이물질 펩타이드의 분해에 역할을 할 수 있다.[16]20S 프로테아좀은 ATP 분해 효소가 없는 7량체 구조인 11S 조절 입자라는 두 번째 유형의 조절 입자와도 결합할 수 있으며, 짧은 펩타이드는 분해할 수 있지만 완전한 단백질은 분해할 수 없다.[41] 이는 복합체가 더 큰 기질을 펼칠 수 없기 때문으로 추정된다. 이 구조는 PA28, REG 또는 PA26으로도 알려져 있다.[41] 이 구조가 하위 단위체의 C-말단 꼬리를 통해 핵심 입자에 결합하고 20S 게이트를 열기 위해 α-고리의 입체구조 변화를 유도하는 메커니즘은 19S 입자와 유사한 메커니즘을 제안한다.[42] 11S 입자의 발현은 감마 인터페론에 의해 유도되며, 면역 프로테아좀 β 소단위와 함께 주요 조직 적합성 복합체에 결합하는 펩타이드를 생성하는 역할을 한다.[16]
26S 프로테아좀은 프로테아제 활성을 갖는 원통형의 20S 프로테아좀에 그 덮개 역할을 하는 19S 복합체가 2개 결합한 것으로, "덤벨형"이라고도 한다. 그에 비해 20S 프로테아좀의 양쪽에 11S 복합체가 각각 결합한 것은 "풋볼형"이라고 불린다.
IFNγ가 생성되면 이에 응답하여 PA28(proteasome activator) 복합체를 유도한다. 이 PA28 복합체는 19S 복합체와 마찬가지로 20S 프로테아좀과 결합한다. 분해된 펩티드 조각의 배출을 촉진하는 것이 그 역할로 생각되며, 19S 복합체-20S 면역 프로테아좀-PA28 복합체의 3가지에 의해 형성된 프로테아좀(하이브리드 프로테아좀)은 MHC 클래스 I에 의해 항원 제시될 때 적합한 펩티드 조각을 생성한다고 생각된다.
2. 4. PA200/Blm10
주어진 원본 소스에는 PA200/Blm10에 대한 내용이 없으므로, 해당 섹션은 작성할 수 없다. (이 부분은 이전 답변과 동일하며, 변경할 내용이 없음)3. 면역 프로테아좀
주요 조직 적합성 복합체 결합에 최적의 크기와 조성을 가진 펩타이드를 주로 생성하는 특수 복합체는 인터페론 감마에 의해 유전자 발현이 유도되는 단백질로 구성된다. 면역 반응 동안 발현이 증가하는 이러한 단백질에는 MHC 리간드의 생성을 조절하는 것으로 알려진 주요 생물학적 역할을 하는 11S 조절 입자와 기질 특이성이 변경된 β1i, β2i, β5i라고 하는 특수 β 서브유닛이 포함된다. 특수 β 서브유닛으로 형성된 복합체는 ''면역 프로테아좀''으로 알려져 있다.[16] 20S 프로테아좀은 ATP 분해 효소가 없는 7량체 구조인 11S 조절 입자와 결합할 수 있으며, 짧은 펩타이드는 분해할 수 있지만 완전한 단백질은 분해할 수 없다. 이는 복합체가 더 큰 기질을 펼칠 수 없기 때문으로 추정된다. 11S 입자의 발현은 감마 인터페론에 의해 유도되며, 면역 프로테아좀 β 소단위와 함께 주요 조직 적합성 복합체에 결합하는 펩타이드를 생성하는 역할을 한다.[16]
IFNγ는 바이러스 감염 등에 의해 생성되며, 이에 대한 응답으로 β1i, β2i, β5i의 3종류의 서브유닛이 유도된다. 이들은 20S 프로테아좀의 β1, β2, β5와 교체되며, 프로테아제 활성 부위가 완전히 바뀌어 면역 프로테아좀이라고 불리는 프로테아좀을 형성한다.
또한 IFNγ는 PA28(proteasome activator) 복합체를 유도한다. 이 PA28 복합체는 19S 복합체와 마찬가지로 20S 프로테아좀과 결합한다. 분해된 펩타이드 조각의 배출을 촉진하는 것이 그 역할로 생각되며, 19S 복합체-20S 면역 프로테아좀-PA28 복합체의 3가지에 의해 형성된 프로테아좀(하이브리드 프로테아좀)은 MHC 클래스 I에 의해 항원 제시될 때 적합한 펩티드 조각을 생성한다고 생각된다.
4. 단백질 분해 기작
프로테아좀은 엔도프로테아제로 기능한다.[65][66][67][68] 20S 코어 입자의 β 소단위체에 의한 단백질 분해 기작은 트레오닌 의존적인 친핵성 공격을 통해 이루어진다. 이 기작은 반응성 트레오닌 수산기의 탈양성자화를 위해 관련된 물 분자에 의존할 수 있다. 분해는 두 개의 β 링 결합으로 형성된 중앙 챔버 내에서 발생하며, 기질을 짧은 폴리펩타이드로 분해한다. 이 폴리펩타이드는 보통 7~9개의 잔기로 구성되지만, 유기체와 기질에 따라 4~25개 잔기 범위를 가질 수 있다. 생성물 길이를 결정하는 생화학적 기작은 완전히 밝혀지지 않았다.[69] 세 개의 촉매 β 소단위체는 공통 기작을 가지지만, 키모트립신 유사, 트립신 유사, 펩티딜-글루타밀 펩타이드-가수분해(PHGH) 유사로 간주되는 약간 다른 기질 특이성을 갖는다. 이러한 특이성 변화는 각 소단위체의 활성 부위 근처 국소 잔기와 원자 간 접촉 결과이다. 각 촉매 β 소단위체는 단백질 분해에 필요한 보존된 라이신 잔기를 갖는다.[21]
프로테아좀은 일반적으로 매우 짧은 펩타이드 조각을 생성하지만, 어떤 경우에는 이 생성물이 생물학적 활성을 띠며 기능적 분자이기도 하다. 전사 인자 중 특정 유전자 발현을 조절하는 일부 전사 인자(예: 포유류 복합체 NF-κB의 한 구성 요소)는 비활성 전구체로 합성되며, 유비퀴틴화와 후속적인 프로테아좀 분해를 통해 활성 형태로 변환된다.
단백질 분해는 항상 유비퀴틴 표지를 필요로 하는 것은 아니다. 오르니틴 탈카르복실화 효소(ODC)가 대표적인 예외이다.[141] 세포 주기 조절에 관여하는 p53에도 유비퀴틴 비의존적 분해 경로가 존재하지만, 이 분자는 유비퀴틴 의존적 분해도 일어난다.[142]
4. 1. 유비퀴틴-프로테아좀 경로
세포 내 단백질 분해는 이전에는 주로 리소좀에 의존하는 것으로 알려졌다. 리소좀은 산성 환경에서 프로테아제를 통해 외부 단백질과 손상된 세포 소기관을 분해하는 세포 소기관이다.[1] 그러나 1977년 망상 적혈구에서 ATP 의존적 단백질 분해에 대한 조셉 에틀링거(Joseph Etlinger)와 알프레드 L. 골드버그(Alfred L. Goldberg)의 연구는 리소좀 외 다른 분해 메커니즘이 존재함을 시사했다.[5] 1978년에는 여러 단백질 사슬로 구성된 새로운 프로테아제가 발견되었다.[6] 히스톤 변형 연구를 통해, 히스톤의 라이신 측쇄와 유비퀴틴의 C-말단 글리신 잔기 사이 결합으로 히스톤이 공유 결합 변형을 일으킨다는 사실이 확인되었다.[7] 이후, ATP 의존적 프로테올리시스 인자 1(APF-1)이 유비퀴틴과 동일하다는 것이 밝혀졌다.[8] 이 시스템의 단백질 분해 활성은 셔윈 윌크(Sherwin Wilk)와 마리온 오를로프스키(Marion Orlowski)에 의해 다중 촉매 단백질 분해 복합체로 분리되었다.[9] 이후, 유비퀴틴 의존적 단백질 분해를 담당하는 ATP 의존적 단백질 분해 복합체가 발견되어 26S 프로테아좀으로 명명되었다.[10][11]테크니온에서 아브람 허쉬코(Avram Hershko)와 아론 체하노버(Aaron Ciechanover)가 1970년대 후반과 1980년대 초에 유비퀴틴 프로테아좀 시스템 발견의 초기 연구를 수행했다. 어윈 로즈(Irwin Rose)는 핵심적인 개념을 제공했지만, 이후 자신의 역할은 축소했다.[12] 이들은 2004년 노벨 화학상을 공동 수상했다.[4]
1980년대 중반 전자 현미경 데이터로 프로테아좀의 적층 링 구조가 밝혀졌고,[13] 1994년 X선 결정학으로 프로테아좀 코어 입자의 첫 번째 구조가 밝혀졌다.[14] 2018년에는 극저온 전자 현미경을 통해 인간 26S 프로테아좀 홀로효소의 원자 구조가 밝혀져 기질 인식, 탈유비퀴틴화, 펼쳐짐 및 분해 메커니즘이 규명되었다.[15]
단백질은 3가지 효소의 반응을 통해 라이신 잔기의 공유 결합적 변형으로 프로테아좀에 의해 분해된다. 유비퀴틴 활성화 효소(E1)는 ATP를 가수분해하고 유비퀴틴 분자를 아데닐화한다. 이후 두 번째 유비퀴틴의 아데닐화와 함께 E1의 활성 부위 시스테인 잔기로 옮겨진다.[48] 아데닐화된 유비퀴틴은 유비퀴틴 결합 효소(E2)의 시스테인으로 옮겨진다. 유비퀴틴 연결 효소(E3)는 유비퀴틴화될 단백질을 인식하고 E2에서 표적 단백질로 유비퀴틴 이동을 촉매한다. 프로테아좀 뚜껑이 인식하기 전, 표적 단백질은 최소 4개의 유비퀴틴 단량체(폴리유비퀴틴 사슬)로 표시되어야 한다.[49] E3가 이 시스템에 기질 특이성을 부여한다.[50] 인간에게는 많은 수의 서로 다른 E3 효소가 존재하여 유비퀴틴 프로테아좀 시스템에 대한 표적이 많음을 나타낸다.
폴리유비퀴틴화된 단백질이 프로테아좀으로 표적화되는 메커니즘은 완전히 이해되지 않았다. 유비퀴틴 수용체가 초기 기질 표적화 및 결합을 위해 탈유비퀴틴화 효소 Rpn11과 조율될 수 있다.[15] 유비퀴틴 수용체 단백질은 N-말단 유비퀴틴 유사(UBL) 도메인과 유비퀴틴 관련(UBA) 도메인을 갖는다. UBL 도메인은 19S 프로테아좀 캡에 의해 인식되고 UBA 도메인은 3중 나선 다발을 통해 유비퀴틴에 결합한다. 이러한 수용체 단백질은 폴리유비퀴틴화된 단백질을 프로테아좀으로 안내할 수 있지만, 자세한 기작은 불분명하다.[51]
유비퀴틴은 76개의 아미노산으로 구성되며, 고도로 보존된 서열을 가지며 모든 진핵생물에서 발견된다.[52] 진핵생물에서 유비퀴틴을 암호화하는 유전자는 탠덤 반복으로 배열되어 세포에 충분한 유비퀴틴을 생성한다. 유비퀴틴은 가장 느리게 진화하는 단백질로 제안되었다.[53] 유비퀴틴은 7개의 라이신 잔기를 포함하여 다양한 유형의 폴리유비퀴틴 사슬을 생성한다.[54] 각 추가 유비퀴틴이 이전 유비퀴틴의 라이신 48에 연결된 사슬은 프로테아좀 표적화에 역할을 하며, 다른 사슬은 다른 과정에 관여한다.[55][56]
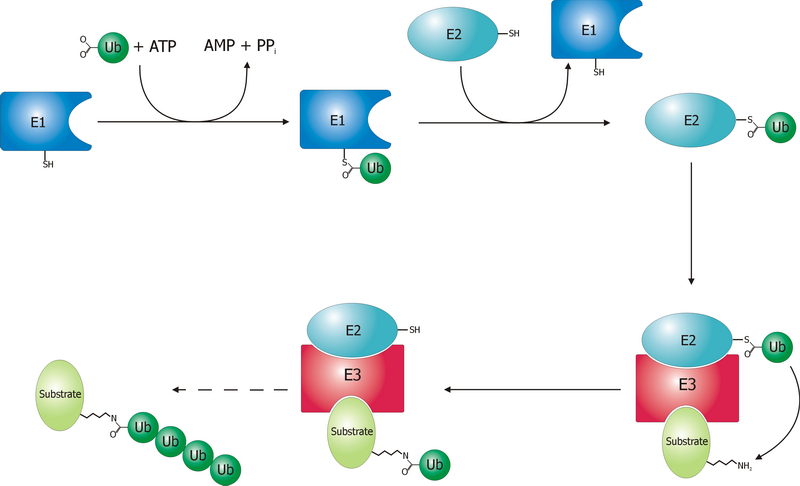
# 표적단백질이 유비퀴틴활성화 효소(E1), 결합효소(E2), 전이효소(E3)에 의해 유비퀴틴화(Ubiquitination) 된다.
# 표적단백질에 결합한 유비퀴틴 사슬이 19S복합체에 결합한다.
# 표적단백질로부터 유비퀴틴사슬이 잘려 떨어진다. 잘려 떨어진 유비퀴틴은 재이용된다.
# 표적단백질의 입체구조가 풀려(unfolding) 20S프로테아솜 속으로 들어간다.
# β-ring내부의 프로테아제활성에 의해 표적단백질이 분해된다.
프로테아좀에 표적화된 단백질에 접합된 유비퀴틴 사슬은 Rpn11, Ubp6/USP14 및 UCH37에 의해 제거된다. 이 과정은 유비퀴틴을 재활용하여 세포 내 유비퀴틴 저장고를 유지한다.[56] Rpn11은 19S 조절 입자의 고유 소단위체이며 26S 프로테아좀 기능에 필수적이다. Rpn11의 DUB 활성은 프로테아좀에서 향상된다. Rpn11이 단백질 기질에서 유비퀴틴 사슬을 제거하는 방법은 EB 형태의 기질 결합 인간 프로테아좀의 원자 구조에 의해 포착되었다.[15] 이 구조는 DUB 활성이 프로테아좀 AAA-ATPase에 의한 기질 인지에 결합되는 방식을 보여준다. USP14와 UCH37은 항상 프로테아좀과 연관되지 않는 DUB이다. 세포에서 프로테아좀의 약 10-40%가 USP14와 연관된다. Ubp6/USP14와 UCH37은 프로테아좀에 의해 활성화되며 단독으로는 낮은 DUB 활성을 나타낸다. USP14는 DUB 활성과 프로테아좀의 형태 변화 경로를 유도하여 프로테아좀 기능을 억제하며, AAA-ATPase로의 기질 삽입을 직접 금지한다.[57] USP14는 Rpn11과 경쟁하고 AAA-ATPase 상태를 재프로그래밍하여 프로테아좀 기능을 조절하며, 이는 DUB로서는 예상 밖의 일이다.[57] 이러한 관찰은 프로테아좀 조절이 형태적 상태의 동적 변화에 의존할 수 있음을 시사한다.
단백질이 유비퀴틴화되면, ATP 의존적 결합 단계에서 19S 조절 입자에 의해 인식된다.[15][27] 기질 단백질은 20S 소단위체의 내부로 들어가야 한다. 20S 입자의 중앙 채널은 좁고 α 링 소단위체의 N-말단 꼬리에 의해 게이트가 형성되어, 기질은 코어에 들어가기 전 부분적으로 펼쳐져야 한다.[15] 펼쳐진 기질이 코어로 들어가는 것을 '전위'라 하며, 탈유비퀴틴화 후에 발생한다.[15][27] 기질이 탈유비퀴틴화되고 펼쳐지는 순서는 불분명하다.[58] 어떤 과정이 속도 제한 단계인지는 기질에 따라 다르다. 일부 단백질은 펼쳐짐 과정이, 다른 단백질은 탈유비퀴틴화가 가장 느린 단계이다.[26] 탈유비퀴틴화 호환 상태에서 기질이 결합된 26S 프로테아좀의 원자 구조에 따르면, 전위 전에 기질이 약 20개의 아미노산 잔기만큼 펼쳐져야 한다.[15] 이황화 결합과 같은 비국소적 상호작용은 분해를 억제하기에 충분하다.[59] 단백질 말단 또는 내부에 충분한 크기의 본질적으로 무질서한 단백질 세그먼트가 존재하면 분해를 촉진한다.[60][61]
α 소단위체에 의해 형성된 게이트는 약 4개 이상의 잔기로 구성된 펩타이드가 20S 입자 내부로 들어가는 것을 막는다. 초기 인식 단계 전에 결합된 ATP 분자는 전위 전에 가수분해된다. 기질 펼쳐짐에는 에너지가 필요하지만, 전위에는 필요하지 않다.[26][27] 조립된 26S 프로테아좀은 비가수분해성 ATP 유사체 존재 시 펼쳐진 단백질을 분해할 수 있지만, 접힌 단백질은 분해할 수 없어 ATP 가수분해 에너지가 기질 펼쳐짐에 사용됨을 나타낸다.[26] 펼쳐진 기질이 열린 게이트를 통과하는 것은 19S 캡이 ATP 결합 상태일 때 촉진 확산을 통해 발생한다.[62]
구상 단백질의 펼쳐짐 메커니즘은 일반적이지만, 아미노산 서열에 따라 달라진다. 글리신과 알라닌이 교대로 나타나는 긴 서열은 기질 펼쳐짐을 억제하고 프로테아좀 분해 효율을 감소시킨다. 이는 ATP 가수분해와 펼쳐짐 단계의 분리로 부분 분해 부산물 방출을 초래한다.[63] 글리신-알라닌 반복 서열은 실크 피브로인에서 발견되며, 이 서열을 가진 엡스타인-바 바이러스 유전자 산물은 프로테아좀을 정지시켜 주요 조직 적합성 복합체에서의 항원 제시를 막아 바이러스 증식을 돕는다.[64]
4. 2. 유비퀴틴 비의존적 분해
대부분의 프로테아좀 기질은 분해되기 전에 유비퀴틴화되어야 하지만, 이러한 일반적인 규칙에는 몇 가지 예외가 있다. 특히 프로테아좀이 단백질의 번역 후 가공에서 정상적인 역할을 수행할 때 더욱 그렇다. p105를 내부 단백질 분해를 통해 p50으로 가공하여 NF-κB를 활성화하는 것이 주요한 예이다.[70] 본질적으로 무질서한 영역 때문에 불안정하다고 추정되는 일부 단백질은 유비퀴틴 비의존적 방식으로 분해된다.[72] 유비퀴틴 비의존적 프로테아좀 기질의 가장 잘 알려진 예는 효소 오르니틴 탈카복실화 효소이다.[73] p53과 같은 주요 세포 주기 조절인자를 표적으로 하는 유비퀴틴 비의존적 메커니즘도 보고되었지만, p53은 유비퀴틴 의존적 분해에도 적용된다.[74] 마지막으로, 구조적으로 비정상적이거나, 잘못 접히거나, 고도로 산화된 단백질도 세포 스트레스 조건에서 유비퀴틴 비의존적이고 19S 비의존적 분해를 받는다.[75]5. 프로테아좀의 생물학적 의의
세포 주기 진행은 특정 사이클린에 의해 활성화되는 사이클린 의존성 키나아제(CDK)의 순차적인 작용에 의해 제어되며, 이는 세포 주기의 단계를 구분한다. 유사 분열 사이클린은 세포 내에서 불과 몇 분 동안만 유지되는데, 이는 모든 세포 내 단백질 중에서 가장 짧은 수명을 가진 단백질 중 하나이다.[1] CDK-사이클린 복합체가 기능을 수행한 후, 관련된 사이클린은 폴리유비퀴틴화되어 프로테아좀에 의해 파괴되며, 이는 세포 주기의 방향성을 제공한다. 특히, 유사 분열 종료에는 조절 성분인 사이클린 B가 유사 분열 촉진 인자 복합체에서 프로테아좀 의존적으로 해리되는 것이 필요하다.[78] 척추동물 세포에서, 방추체 점검 지점에 의해 이탈이 지연됨에도 불구하고, 유사 분열 점검 지점을 통과하는 "슬립" 현상이 발생하여 조기 M기 종료가 발생할 수 있다.[79]
G1기와 S기 사이의 제한점 이후와 같은 이전의 세포 주기 점검 지점도, E3 유비퀴틴 연결 효소인 후기 촉진 복합체(APC)에 의해 유비퀴틴화가 촉진되는 사이클린 A의 프로테아좀 분해를 유사하게 포함한다.[80] APC와 Skp1/Cul1/F-box 단백질 복합체 (SCF 복합체)는 사이클린 분해 및 점검 지점 제어의 두 가지 주요 조절자이며, SCF 자체는 G1-S 전환 전에 SCF 활성을 방지하는 어댑터 단백질 Skp2의 유비퀴틴화를 통해 APC에 의해 조절된다.[81]
19S 입자의 개별 구성 요소는 자체 조절 역할을 한다. 최근에 확인된 종양 유전자인 강키린은 사이클린 의존성 키나아제 CDK4에 강력하게 결합하며 유비퀴틴 연결 효소 MDM2에 대한 친화성을 통해 유비퀴틴화된 p53을 인식하는 데 중요한 역할을 하는 19S 서브 구성 요소 중 하나이다. 강키린은 항세포 사멸 작용을 하며, 간세포 암종과 같은 일부 종양 세포 유형에서 과발현되는 것으로 나타났다.[82]
진핵생물과 마찬가지로, 일부 고세균도 프로테아좀을 사용하여 세포 주기를 제어하며, 특히 ESCRT-III 매개 세포 분열을 제어한다.[83]
식물에서 식물 생장의 방향과 굴성을 지시하는 옥신 또는 식물 호르몬에 의한 신호 전달은, 프로테아좀 분해를 위해 Aux/IAA 단백질로 알려진 일련의 전사 인자 억제 물질의 표적화를 유도한다. 이 단백질은 옥신 수용체 TIR1과 복합체를 이룬 SCFTIR1 또는 SCF에 의해 유비퀴틴화된다. Aux/IAA 단백질의 분해는 옥신 반응 인자(ARF) 계열의 전사 인자를 억제 해제하고 ARF가 지시하는 유전자 발현을 유도한다.[84] ARF 활성화의 세포 결과는 식물의 유형 및 발달 단계에 따라 다르지만, 뿌리와 잎맥의 생장을 지시하는 데 관여한다. ARF 억제 해제에 대한 특이적인 반응은 개별 ARF 및 Aux/IAA 단백질의 쌍을 이루는 특이성에 의해 매개되는 것으로 생각된다.[85]
내부 및 외부의 세포 신호 전달 모두 세포 자멸사 또는 프로그램된 세포 사멸을 유도할 수 있다. 그 결과 세포 구성 요소의 해체는 주로 카스파제로 알려진 특수 단백질 분해 효소에 의해 수행되지만, 프로테아좀 또한 세포 자멸 과정에서 중요하고 다양한 역할을 수행한다. 이 과정에서 프로테아좀의 관여는 세포 자멸사보다 훨씬 전에 관찰되는 단백질 유비퀴틴화와 E1, E2 및 E3 효소의 증가로 나타난다.[86][87][88] 세포 자멸사 동안, 핵에 위치한 프로테아좀은 세포 자멸사의 특징인 외부 막 블레브로 이동하는 것이 관찰되었다.[89]
프로테아좀 억제는 다른 세포 유형에서 세포 자멸사 유도에 서로 다른 영향을 미친다. 일반적으로 프로테아좀은 세포 자멸사에 필요하지 않지만, 이를 억제하는 것은 연구된 대부분의 세포 유형에서 세포 자멸사를 촉진한다. 세포 자멸사는 세포 성장 촉진 단백질의 조절된 분해를 방해함으로써 매개된다.[102] 그러나 일부 세포주, 특히 G0기의 휴지기 및 세포 분화된 세포, 예를 들어 흉선 세포 및 신경 세포의 일차 배양은 프로테아좀 억제제에 노출되었을 때 세포 자멸사를 겪지 못하게 된다. 이러한 효과의 기전은 불분명하지만, 휴지 상태의 세포에 특이적이거나, 세포 자멸사를 촉진하는 키나아제 JNK의 차등 활성화의 결과일 것으로 추정된다.[90] 빠르게 분열하는 세포에서 프로테아좀 억제제가 세포 자멸사를 유도하는 능력은 최근 개발된 여러 화학 요법 제제, 예를 들어 보르테조미브와 Salinosporamide A|살리노스포라미드 A영어에서 활용되었다.
세포 스트레스, 예를 들어 감염, 열 충격, 또는 산화 손상에 대한 반응으로, 잘못 접히거나 펼쳐진 단백질을 식별하고 프로테아좀 분해를 목표로 하는 열 충격 단백질이 발현된다. Hsp27과 Hsp90 (샤페론 단백질)은 유비퀴틴-프로테아좀 시스템의 활성을 증가시키는 데 관여하지만, 이 과정에 직접적으로 참여하지는 않는다.[91] 반면에 Hsp70은 잘못 접힌 단백질 표면의 노출된 소수성 패치에 결합하여 CHIP과 같은 E3 유비퀴틴 연결 효소를 모집하여 프로테아좀 분해를 위해 단백질에 태그를 붙인다.[92] CHIP 단백질(Hsp70-상호 작용 단백질의 카르복시 말단) 자체는 E3 효소 CHIP과 E2 결합 파트너 간의 상호 작용 억제를 통해 조절된다.[93]
프로테아좀 시스템을 통해 산화적으로 손상된 단백질의 분해를 촉진하는 유사한 메커니즘이 존재한다. 특히, 핵에 국한된 프로테아좀은 PARP에 의해 조절되며 부적절하게 산화된 히스톤을 활발하게 분해한다.[94] 종종 세포 내에서 큰 비정질 응집체를 형성하는 산화 단백질은 19S 조절 캡 없이 20S 코어 입자에 의해 직접 분해될 수 있으며, ATP 가수분해 또는 유비퀴틴으로의 태깅이 필요하지 않다.[75] 그러나 높은 수준의 산화 손상은 단백질 조각 간의 가교 결합 정도를 증가시켜 응집체가 단백질 분해에 저항하게 만든다. 이러한 고도로 산화된 응집체의 더 많은 수와 크기는 노화와 관련이 있다.[95]
유비퀴틴 프로테아좀 시스템의 조절 장애는 여러 신경 질환에 기여할 수 있다. 이는 성상 세포종과 같은 뇌종양으로 이어질 수 있다.[96] 파킨슨병 및 알츠하이머병과 같이 잘못 접힌 단백질의 응집을 공통 특징으로 공유하는 일부 후기 발병 신경퇴행성 질환에서, 잘못 접힌 단백질의 크고 불용성 응집체가 형성되어 아직 잘 이해되지 않은 메커니즘을 통해 신경 독성을 초래할 수 있다. 감소된 프로테아좀 활성은 파킨슨병에서 응집 및 루이 소체 형성의 원인으로 제시되었다.[97] 이러한 가설은 파킨슨병의 효모 모델이 낮은 프로테아좀 활성 조건에서 루이 소체의 주요 단백질 성분인 α-시누클레인의 독성에 더 취약하다는 관찰에 의해 뒷받침된다.[98] 손상된 프로테아좀 활성은 자폐 스펙트럼 장애와 같은 인지 장애, 봉입체 근병증과 같은 근육 및 신경 질환의 근본 원인이 될 수 있다.[96]
단백질 분해 효소는 적응 면역 체계의 기능에 직접적이지만 중요한 역할을 한다. 펩타이드 항원은 주요 조직 적합성 복합체 I형(MHC) 단백질에 의해 항원 제시 세포 표면에 표시된다. 이러한 펩타이드는 침입하는 병원체에서 유래된 단백질의 단백질 분해 효소에 의한 분해 산물이다. 구성적으로 발현되는 단백질 분해 효소도 이 과정에 참여할 수 있지만, 인터페론 감마에 의해 유전자 발현이 유도되는 단백질로 구성된 특수 복합체는 MHC 결합에 최적의 크기와 조성을 가진 펩타이드를 주로 생성한다. 면역 반응 동안 발현이 증가하는 이러한 단백질에는 MHC 리간드의 생성을 조절하는 것으로 알려진 주요 생물학적 역할을 하는 11S 조절 입자와 기질 특이성이 변경된 β1i, β2i, β5i라고 하는 특수 β 서브유닛이 포함된다. 특수 β 서브유닛으로 형성된 복합체는 ''면역 단백질 분해 효소''로 알려져 있다.[16] 또 다른 β5i 변이체 서브유닛인 β5t는 흉선에서 발현되어 기능이 아직 불분명한 흉선 특이적 "흉선 단백질 분해 효소"를 생성한다.[99]
MHC I형 리간드 결합의 강도는 리간드 C-말단의 조성에 따라 달라지며, 펩타이드는 수소 결합과 MHC 표면의 "B 포켓"이라고 하는 영역과의 밀접한 접촉으로 결합한다. 많은 MHC I형 대립 유전자는 소수성 C-말단 잔기를 선호하며, 면역 단백질 분해 효소 복합체는 소수성 C-말단을 생성할 가능성이 더 높다.[100]
단백질 분해 효소의 활성은 NF-κB의 활성화 형태 생성에 관여하며, 세포 자멸사 억제 및 염증 유발 사이토카인 발현 조절과 관련이 있어 염증성 및 자가 면역 질환과 관련이 있다. 단백질 분해 효소 활성의 증가된 수준은 질병 활동과 상관관계가 있으며 전신성 홍반성 루푸스 및 류마티스 관절염을 포함한 자가 면역 질환에 관여한다.[16]
프로테아좀은 또한 항체에 결합된 바이러스 입자의 세포 내 항체 매개 단백질 분해에 관여한다. 이 중화 경로에서 TRIM21 (삼분할 모티프 계열의 단백질)은 면역글로불린 G와 결합하여 바이러스 입자를 단백질 분해 효소로 보내 분해시킨다.[101]
프로테아좀과 마찬가지로 세포 내 단백질 분해를 담당하는 것으로 리소좀이 알려져 있지만, 리소좀은 단백질을 분해하여 다른 새로운 단백질을 합성하기 위한 재료를 공급하는 것을 목적으로 하는 데 비해, 프로테아좀은 목적 단백질을 특이적으로 분해하여 세포 내에서 제거하는 것을 목적으로 한다.
6. 프로테아좀 관련 질병

보르테조밉(VELCADE)은 26S프로테아좀을 억제하는 약물로, 혈액암의 일종인 다발성골수종 치료에 효과가 있어 항암제로 사용된다.[145] 세포는 감염, 열 충격, 산화 손상 등 스트레스에 반응하여 열 충격 단백질을 발현하는데, 이는 잘못 접히거나 펼쳐진 단백질을 식별하고 프로테아좀 분해를 유도한다. Hsp27, Hsp90과 같은 샤페론 단백질은 유비퀴틴-프로테아좀 시스템 활성을 증가시키지만, 직접 관여하지는 않는다.[91] 반면 Hsp70은 잘못 접힌 단백질의 소수성 부분에 결합, CHIP 같은 E3 유비퀴틴 연결 효소를 모집하여 프로테아좀 분해를 위한 표식을 붙인다.[92]
산화적으로 손상된 단백질은 프로테아좀 시스템을 통해 분해 촉진된다. 핵 내 프로테아좀은 PARP에 의해 조절되며 산화된 히스톤을 분해한다.[94] 산화 단백질은 종종 큰 응집체를 형성하며, 19S 조절 캡 없이 20S 코어 입자에 의해 직접 분해될 수 있다(ATP 가수분해, 유비퀴틴 표식 불필요).[75] 그러나 심한 산화 손상은 단백질 조각 간 교차 결합을 증가시켜 응집체의 단백질 분해 저항성을 높인다. 이러한 산화 응집체는 노화와 관련이 깊다.[95]
유비퀴틴 프로테아좀 시스템 조절 장애는 성상 세포종 등 여러 신경 질환 및 뇌종양을 유발할 수 있다.[96] 파킨슨병, 알츠하이머병과 같이 잘못 접힌 단백질 응집이 특징인 신경퇴행성 질환에서는, 크고 불용성인 응집체가 형성되어 신경 독성을 일으킨다. 프로테아좀 활성 감소는 파킨슨병에서 응집 및 루이 소체 형성의 원인으로 제시되었다.[97] 손상된 프로테아좀 활성은 자폐 스펙트럼 장애, 봉입체 근병증 등의 질환 원인이 될 수 있다.[96]
프로테아좀과 그 하위 단위체는 (1) 복합체 조립 손상/기능 장애가 특정 질병과 관련, (2) 치료 약물 표적으로 활용 가능하여 임상적으로 중요하다. 최근에는 프로테아좀을 진단 표지 및 전략 개발에 활용하려는 노력이 진행 중이다.
프로테아좀은 유비퀴틴-프로테아좀 시스템(UPS) 및 세포 내 단백질 품질 관리(PQC)의 핵심이다. 단백질 유비퀴틴화와 프로테아좀에 의한 단백질 분해는 세포 주기, 세포 성장/분화, 유전자 전사, 신호 전달, 세포 사멸 조절에 중요하다.[119] 프로테아좀 결함은 단백질 분해 활성 감소, 손상/잘못 접힌 단백질 축적을 유발하여 신경 퇴행성 질환,[120][121] 심혈관 질환,[122][123][124] 염증 반응, 자가면역 질환,[125] 악성 종양 유발 DNA 손상 반응에 기여한다.[126]
UPS 결함은 알츠하이머병,[127] 파킨슨병[128], 픽병,[129] 근위축성 측삭 경화증(ALS),[129] 헌팅턴병,[128] 크로이츠펠트-야코프병,[130] 운동 신경 질환, 폴리글루타민(PolyQ) 질환, 근이영양증,[131] 치매 관련 신경 퇴행성/근육 퇴행성 장애에 관여한다.[132] 프로테아좀은 심장 단백질 항상성 유지로 심장 허혈 손상,[133] 심실 비대[134], 심부전에 중요하다.[135] UPS는 암 발달 관련 신호에 대한 암세포 반응에 중요하며, 전사 인자(p53, c-jun, c-Fos, NF-κB, c-Myc, HIF-1α, MATα2, STAT3, 스테롤 조절 요소 결합 단백질, 안드로겐 수용체) 분해에 의한 유전자 발현을 제어하여 다양한 악성 종양 발생에 관여한다.[136] UPS는 종양 억제 유전자 산물(APC, Rb, VHL)과 원종양 유전자(Raf, Myc, Myb, Rel, Src, Mos, ABL) 분해를 조절한다. 또한, 염증성 사이토카인(TNF-α, IL-β, IL-8, 부착 분자(ICAM-1, VCAM-1, P-selectin), 프로스타글란딘, NO) 발현을 조절하는 NF-κB 활성화에 관여하여 염증 반응을 조절하고,[125] 백혈구 증식 조절에도 역할[137]을 한다. 전신성 홍반성 루푸스(SLE), 쇼그렌 증후군, 류마티스 관절염(RA) 환자는 순환 프로테아좀을 나타낸다.[138]
7. 프로테아좀 억제제
프로테아좀 억제제는 세포 배양에서 효과적인 항-종양 활성을 보이며, 성장 촉진 세포 주기 단백질의 조절된 분해를 방해하여 세포자멸사를 유도한다.[102] 종양 세포에서 세포자멸사를 선택적으로 유도하는 이러한 접근 방식은 동물 모델 및 인간 임상 시험에서 효과적인 것으로 입증되었다.
락타시스틴은 ''방선균'' 세균에 의해 합성된 천연 생성물로, 최초로 발견된 비펩티드 프로테아좀 억제제이며,[103] 생화학 및 세포 생물학 연구 도구로 널리 사용된다. 락타시스틴은 프로테아좀의 촉매 β 서브유닛, 특히 프로테아좀의 키모트립신 유사 활성을 담당하는 β5 서브유닛의 아미노 말단 트레오닌을 공유 결합 방식으로 변형시킨다. 이러한 발견은 프로테아좀을 메커니즘적으로 새로운 종류의 프로테아제, 즉 아미노 말단 트레오닌 프로테아제로 확립하는 데 기여했다.
보르테조밉 (붕소화 MG132)은 밀레니엄 제약에서 개발하여 Velcade라는 이름으로 판매되는 분자로, 화학요법제로 임상 사용에 도달한 최초의 프로테아좀 억제제이다.[104] 다발성 골수종 치료에 사용된다.[105] 특히 다발성 골수종은 성공적인 화학 요법에 반응하여 정상 수준으로 감소하는 혈청의 프로테아좀 유래 펩타이드 수치가 증가하는 것으로 관찰되었다.[106] 동물 연구에 따르면 췌장암에서도 임상적으로 유의미한 효과를 보일 수 있다.[107][108] 보르테조밉의 효과를 검토하기 위한 전임상 및 초기 임상 연구가 다른 B 세포 관련 암,[109] 특히 일부 유형의 비호지킨 림프종 치료에 대해 시작되었다.[110] 임상 결과는 또한 B 세포 급성 림프구성 백혈병에 대한 화학 요법과 결합된 프로테아좀 억제제의 사용을 정당화하는 것으로 보인다.[111] 프로테아좀 억제제는 글루코코르티코이드에 내성이 있는 일부 유형의 배양 백혈병 세포를 죽일 수 있다.[112] 26S 프로테아좀을 억제하는 보르테조밉은 혈액암의 일종인 다발성 골수종에 효과가 있는 것으로 보고되어 항암제로 임상 응용되고 있다.[143]
Norvir로 판매되는 리토나비어 분자는 프로테아제 억제제로 개발되어 HIV 감염을 표적으로 사용되었다. 그러나 이 약물은 유리 프로테아제뿐만 아니라 프로테아좀도 억제하는 것으로 나타났으며, 구체적으로는 리토나비어에 의해 프로테아좀의 키모트립신 유사 활성이 억제되는 반면, 트립신 유사 활성은 다소 향상된다.[113] 동물 모델 연구에 따르면 신경교종 세포의 성장을 억제하는 효과가 있을 수 있다.[114]
프로테아좀 억제제는 또한 동물 모델에서 자가면역 질환을 치료하는 데 희망을 보여주었다. 예를 들어, 인간 피부 이식을 받은 쥐에 대한 연구에서 프로테아좀 억제제로 치료한 후 건선 병변의 크기가 감소하는 것을 발견했다.[115] 억제제는 또한 설치류 천식 모델에서 긍정적인 효과를 보인다.[116]
프로테아좀의 표지 및 억제는 또한 세포 내 프로테아좀 활성에 대한 ''시험관 내'' 및 ''생체 내'' 연구를 위해 실험실 환경에서도 관심의 대상이다. 가장 일반적으로 사용되는 실험실 억제제는 락타시스틴과 Goldberg 연구소에서 처음 개발한 펩타이드 알데히드 MG132이다. 조립된 프로테아좀의 활성 부위를 구체적으로 표지하기 위해 형광 억제제도 개발되었다.[117]
참조
[1]
서적
Molecular cell biology
https://archive.org/[...]
W.H. Freeman and CO
[2]
논문
Distinct 19 S and 20 S subcomplexes of the 26 S proteasome and their distribution in the nucleus and the cytoplasm
1994-03
[3]
논문
Slipping up: Partial substrate degradation by ATP-dependent proteases
2014-05
[4]
웹사이트
Nobel Prize Awardees in Chemistry, 2004
http://nobelprize.or[...]
Nobel Prize Committee
2006-12-11
[5]
논문
A soluble ATP-dependent proteolytic system responsible for the degradation of abnormal proteins in reticulocytes
1977-01
[6]
논문
A heat-stable polypeptide component of an ATP-dependent proteolytic system from reticulocytes
1978-04
[7]
논문
Isopeptide linkage between nonhistone and histone 2A polypeptides of chromosomal conjugate-protein A24
1977-03
[8]
논문
Early work on the ubiquitin proteasome system, an interview with Aaron Ciechanover. Interview by CDD
2005-09
[9]
논문
Cation-sensitive neutral endopeptidase: isolation and specificity of the bovine pituitary enzyme
1980-11
[10]
논문
Identity of 19S prosome particle with the large multifunctional protease complex of mammalian cells.
1988
[11]
논문
Purification of two high molecular weight proteases from rabbit reticulocyte lysate
http://www.jbc.org/c[...]
1987-06
[12]
논문
Early work on the ubiquitin proteasome system, an interview with Avram Hershko. Interview by CDD
2005-09
[13]
논문
Size and shape of the multicatalytic proteinase from rat skeletal muscle
1986-08
[14]
논문
Crystal structure of the 20S proteasome from the archaeon T. acidophilum at 3.4 A resolution
1995-04
[15]
논문
Cryo-EM structures and dynamics of substrate-engaged human 26S proteasome
2018-11
[16]
논문
The ubiquitin-proteasome system and its role in inflammatory and autoimmune diseases
2006-08
[17]
논문
Docking of the proteasomal ATPases' carboxyl termini in the 20S proteasome's alpha ring opens the gate for substrate entry
2007-09
[18]
웹사이트
MEROPS Family T1
https://www.ebi.ac.u[...]
EMBL-EBI
2019-02-16
[19]
논문
Evidence that pituitary cation-sensitive neutral endopeptidase is a multicatalytic protease complex
1983-03
[20]
논문
The ubiquitin-proteasome system
http://eprints.iisc.[...]
2006-03
[21]
논문
The active sites of the eukaryotic 20 S proteasome and their involvement in subunit precursor processing
1997-10
[22]
논문
Assembly of an Evolutionarily Conserved Alternative Proteasome Isoform in Human Cells
2016-03
[23]
논문
Plasticity in eucaryotic 20S proteasome ring assembly revealed by a subunit deletion in yeast
2004-02
[24]
논문
A multimeric assembly factor controls the formation of alternative 20S proteasomes
2008-03
[25]
논문
An archaebacterial ATPase, homologous to ATPases in the eukaryotic 26 S proteasome, activates protein breakdown by 20 S proteasomes
1999-09
[26]
논문
ATP binding to PAN or the 26S ATPases causes association with the 20S proteasome, gate opening, and translocation of unfolded proteins
2005-12
[27]
논문
ATP binding and ATP hydrolysis play distinct roles in the function of 26S proteasome
2006-10
[28]
논문
A proteasomal ATPase subunit recognizes the polyubiquitin degradation signal
2002-04
[29]
논문
Near-atomic resolution structural model of the yeast 26S proteasome
2012-09
[30]
논문
Complete subunit architecture of the proteasome regulatory particle
2012-02
[31]
논문
Molecular architecture of the 26S proteasome holocomplex determined by an integrative approach
2012-01
[32]
논문
Structural basis for dynamic regulation of the human 26S proteasome
2016-11
[33]
논문
An atomic structure of the human 26S proteasome
2016-09
[34]
논문
Structure of the human 26S proteasome at a resolution of 3.9 Å
2016-07
[35]
논문
Structural mechanism for nucleotide-driven remodeling of the AAA-ATPase unfoldase in the activated human 26S proteasome
2018-04
[36]
논문
Deep classification of a large cryo-EM dataset defines the conformational landscape of the 26S proteasome
2014-04
[37]
논문
Structure of the 26S proteasome with ATP-γS bound provides insights into the mechanism of nucleotide-dependent substrate translocation
2013-04
[38]
논문
Conformational switching of the 26S proteasome enables substrate degradation
2013-07
[39]
논문
Conformational Landscape of the p28-Bound Human Proteasome Regulatory Particle
2017-07
[40]
논문
The axial channel of the proteasome core particle is gated by the Rpt2 ATPase and controls both substrate entry and product release
2001-06
[41]
논문
Proteasome activators.
2011-01-07
[42]
논문
The 1.9 A structure of a proteasome-11S activator complex and implications for proteasome-PAN/PA700 interactions
2005-05
[43]
논문
Proteasome assembly triggers a switch required for active-site maturation
2006-07
[44]
논문
20S proteasome biogenesis
[45]
논문
Molecular mechanisms of proteasome assembly
2009-02
[46]
논문
The catalytic activity of Ubp6 enhances maturation of the proteasomal regulatory particle
2011-06
[47]
논문
Dissection of the assembly pathway of the proteasome lid in Saccharomyces cerevisiae
2010-06
[48]
논문
Ubiquitin-activating enzyme. Mechanism and role in protein-ubiquitin conjugation
1982-03
[49]
논문
Recognition of the polyubiquitin proteolytic signal
2000-01
[50]
논문
Protein interaction analysis of SCF ubiquitin E3 ligase subunits from Arabidopsis
2003-06
[51]
논문
Delivery of ubiquitinated substrates to protein-unfolding machines
2005-08
[52]
논문
The ubiquitin-proteasome system: central modifier of plant signalling
2012-10
[53]
논문
Ubiquitin genes as a paradigm of concerted evolution of tandem repeats
[54]
논문
Polyubiquitin chains: polymeric protein signals
2004-12
[55]
논문
Quantitative proteomics reveals the function of unconventional ubiquitin chains in proteasomal degradation
2009-04
[56]
논문
Ubiquitin in chains
2000-11
[57]
논문
USP14-regulated allostery of the human proteasome by time-resolved cryo-EM
2022-04
[58]
논문
Deubiquitination by proteasome is coordinated with substrate translocation for proteolysis in vivo
2005-07
[59]
논문
Conformational constraints in protein degradation by the 20S proteasome
1995-03
[60]
논문
Defining the geometry of the two-component proteasome degron
2011-03
[61]
논문
Intrinsically disordered segments affect protein half-life in the cell and during evolution
2014-09
[62]
논문
Proteasomes and their associated ATPases: a destructive combination
2006-10
[63]
논문
Glycine-alanine repeats impair proper substrate unfolding by the proteasome
2006-04
[64]
논문
Repeat sequence of Epstein–Barr virus-encoded nuclear antigen 1 protein interrupts proteasome substrate processing
2004-03
[65]
논문
Proteasome from Thermoplasma acidophilum: a threonine protease
1995-04
[66]
논문
Structure and functions of the 20S and 26S proteasomes
1996
[67]
논문
Structure of 20S proteasome from yeast at 2.4 A resolution
1997-04
[68]
논문
Contribution of proteasomal beta-subunits to the cleavage of peptide substrates analyzed with yeast mutants
1998-10
[69]
논문
The 26S proteasome: a molecular machine designed for controlled proteolysis
[70]
논문
Taking a bite: proteasomal protein processing
2002-05
[71]
논문
Productive RUPture: activation of transcription factors by proteasomal processing
2004-11
[72]
논문
20S proteasomes and protein degradation "by default"
2006-08
[73]
논문
Determinants of proteasome recognition of ornithine decarboxylase, a ubiquitin-independent substrate
2003-04
[74]
논문
p53 proteasomal degradation: poly-ubiquitination is not the whole story
2005-08
[75]
논문
Ubiquitin conjugation is not required for the degradation of oxidized proteins by proteasome
2003-01
[76]
논문
A comprehensive view on proteasomal sequences: implications for the evolution of the proteasome
2003-03
[77]
논문
The proteasome
[78]
논문
Cyclin B dissociation from CDK1 precedes its degradation upon MPF inactivation in mitotic extracts of Xenopus laevis embryos
2006-08
[79]
논문
Mitotic checkpoint slippage in humans occurs via cyclin B destruction in the presence of an active checkpoint
2006-06
[80]
논문
Regulation of late G1/S phase transition and APC Cdh1 by reactive oxygen species
2006-06
[81]
논문
Control of the SCF(Skp2-Cks1) ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase
2004-03
[82]
논문
The oncoprotein gankyrin negatively regulates both p53 and RB by enhancing proteasomal degradation
2005-10
[83]
논문
The proteasome controls ESCRT-III–mediated cell division in an archaeon
2020-08-07
[84]
논문
The role of regulated protein degradation in auxin response
[85]
논문
Developmental specificity of auxin response by pairs of ARF and Aux/IAA transcriptional regulators
2005-05
[86]
논문
Coordinated induction of the ubiquitin conjugation pathway accompanies the developmentally programmed death of insect skeletal muscle
1995-04
[87]
논문
Activation of polyubiquitin gene expression during developmentally programmed cell death
1990-10
[88]
논문
Expression of a 26S proteasome ATPase subunit, MS73, in muscles that undergo developmentally programmed cell death, and its control by ecdysteroid hormones in the insect Manduca sexta
1997-01
[89]
논문
Removal of proteasomes from the nucleus and their accumulation in apoptotic blebs during programmed cell death
1996-09
[90]
논문
The role of the ubiquitin-proteasome pathway in apoptosis
1999-04
[91]
논문
Heat shock proteins 27 and 70: anti-apoptotic proteins with tumorigenic properties
2006-11
[92]
논문
The cytoplasmic Hsp70 chaperone machinery subjects misfolded and endoplasmic reticulum import-incompetent proteins to degradation via the ubiquitin-proteasome system
2007-01
[93]
논문
Regulation of the cytoplasmic quality control protein degradation pathway by BAG2
2005-11
[94]
논문
Protein oxidation and proteolysis
[95]
논문
Degradation of oxidized proteins by the 20S proteasome
[96]
논문
The ubiquitin proteasome system in neuropathology
2009-09
[97]
논문
Proteasomal dysfunction in sporadic Parkinson's disease
2006-05
[98]
논문
alpha-Synuclein budding yeast model: toxicity enhanced by impaired proteasome and oxidative stress
[99]
논문
Regulation of CD8+ T cell development by thymus-specific proteasomes
2007-06
[100]
논문
26S proteasomes and immunoproteasomes produce mainly N-extended versions of an antigenic peptide
2001-05
[101]
논문
Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21)
2010-11
[102]
논문
Proteasome inhibitors: a novel class of potent and effective antitumor agents
1999-06
[103]
논문
Inhibition of proteasome activities and subunit-specific amino-terminal threonine modification by lactacystin
1995-05
[104]
웹사이트
United States Food and Drug Administration press release
https://www.fda.gov/[...]
2003-05-13
[105]
논문
Multicenter phase II study of bortezomib in patients with relapsed or refractory mantle cell lymphoma
2006-10
[106]
논문
Circulating proteasome levels are an independent prognostic factor for survival in multiple myeloma
2007-03
[107]
논문
26S proteasome inhibition induces apoptosis and limits growth of human pancreatic cancer
[108]
논문
The proteasome inhibitor bortezomib enhances the activity of docetaxel in orthotopic human pancreatic tumor xenografts
2004-01
[109]
논문
Proteasome inhibitors in the treatment of B-cell malignancies
2002-06
[110]
논문
Phase II clinical experience with the novel proteasome inhibitor bortezomib in patients with indolent non-Hodgkin's lymphoma and mantle cell lymphoma
2005-02
[111]
논문
Bortezomib with chemotherapy is highly active in advanced B-precursor acute lymphoblastic leukemia: Therapeutic Advances in Childhood Leukemia & Lymphoma (TACL) Study
2012-07
[112]
논문
Glucocorticoid and proteasome inhibitor impact on the leukemic lymphoblast: multiple, diverse signals converging on a few key downstream regulators
2012-04
[113]
논문
How an inhibitor of the HIV-I protease modulates proteasome activity
1999-12
[114]
논문
Effects of the proteasome inhibitor ritonavir on glioma growth in vitro and in vivo
2004-02
[115]
논문
Proteasome inhibition reduces superantigen-mediated T cell activation and the severity of psoriasis in a SCID-hu model
2002-03
[116]
논문
Proteasome inhibition: A novel mechanism to combat asthma
1999-08
[117]
논문
A fluorescent broad-spectrum proteasome inhibitor for labeling proteasomes in vitro and in vivo
2006-11
[118]
논문
Perilous journey: a tour of the ubiquitin-proteasome system
2014-06
[119]
논문
New insights into proteasome function: from archaebacteria to drug development
1995-08
[120]
논문
The Ubiquitin–Proteasome System and Molecular Chaperone Deregulation in Alzheimer's Disease
2015-01
[121]
논문
Ubiquitin-proteasome system involvement in Huntington's disease
2014
[122]
논문
Proteotoxicity: an underappreciated pathology in cardiac disease
2014-06
[123]
논문
Targeting the ubiquitin–proteasome system in heart disease: the basis for new therapeutic strategies
2014-12
[124]
논문
Protein quality control and metabolism: bidirectional control in the heart
2015-02
[125]
논문
The I kappa B kinase (IKK) and NF-kappa B: key elements of proinflammatory signalling
2000-02
[126]
논문
Quality control mechanisms in cellular and systemic DNA damage responses
2015-01
[127]
논문
Role of the proteasome in Alzheimer's disease
2000-07
[128]
논문
The role of the ubiquitin-proteasomal pathway in Parkinson's disease and other neurodegenerative disorders
2001-11
[129]
논문
Morphometrical reappraisal of motor neuron system of Pick's disease and amyotrophic lateral sclerosis with dementia
2002-07
[130]
논문
Marked increase in cerebrospinal fluid ubiquitin in Creutzfeldt–Jakob disease
1992-05
[131]
논문
Limb-girdle muscular dystrophy
2003-01
[132]
논문
From neurodegeneration to neurohomeostasis: the role of ubiquitin
2003-03
[133]
논문
The ubiquitin proteasome system and myocardial ischemia
2013-02
[134]
논문
Ubiquitin proteasome dysfunction in human hypertrophic and dilated cardiomyopathies
2010-03
[135]
논문
The ubiquitin-proteasome system in cardiac physiology and pathology
2006-07
[136]
논문
Potential for proteasome inhibition in the treatment of cancer
2003-04
[137]
논문
Regulatory functions of ubiquitination in the immune system
2002-01
[138]
논문
Circulating proteasomes are markers of cell damage and immunologic activity in autoimmune diseases
2002-10
[139]
논문
"Distinct 19S and 20S subcomplexes of the 26S proteasome and their distribution in the nucleus and the cytoplasm."
1994
[140]
논문
"The active sites of the eukaryotic 20 S proteasome and their involvement in subunit precursor processing."
1997
[141]
논문
Repeat sequence of Epstein-Barr virus-encoded nuclear antigen 1 protein interrupts proteasome substrate processing.
2004
[142]
논문
p53 proteasomal degradation: poly-ubiquitination is not the whole story.
2005
[143]
논문
Multicenter phase II study of bortezomib in patients with relapsed or refractory mantle cell lymphoma.
2006
[144]
저널
PACemakers of proteasome core particle assembly
http://www.sciencedi[...]
[145]
논문
Multicenter phase II study of bortezomib in patients with relapsed or refractory mantle cell lymphoma.
2006
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com