고래
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
고래는 육상 포유류에서 진화하여 물속에서 생활하는 해양 포유류로, 긴수염고래류와 이빨고래류로 분류된다. 고래는 폐로 호흡하며, 지느러미와 수평 꼬리지느러미를 사용하여 헤엄친다. 화석 연구와 분자생물학적 연구를 통해 고래가 우제류와 가까운 관계이며, 하마와 가장 가까운 친척으로 밝혀졌다. 고래는 전 세계 바다에 분포하며, 멸종 위협에 직면해 있어 국제적인 보호 노력이 이루어지고 있다.

더 읽어볼만한 페이지
- 1762년 기재된 분류군 - 맥 (동물)
기제목에 속하는 포유류인 맥은 말레이맥, 남아메리카맥, 산악맥, 베어드맥, 카보마니맥 등의 현존 종이 있으며, 짧고 굵은 몸통과 유연한 코와 윗입술을 가진 북아메리카, 남아메리카, 동남아시아 서식 동물로, 서식지 파괴와 사냥으로 일부 종의 개체 수가 감소하고 있다. - 해양 포유류 - 해달
해달은 족제비과에 속하는 해양 포유류로, 뛰어난 수영 실력과 두꺼운 모피를 지녔으며 해양 생태계 유지에 중요한 핵심종이지만, 현재는 기름 유출, 범고래의 포식, 질병 등 다양한 위협에 직면하고 있다. - 해양 포유류 - 바다수달
바다수달은 남아메리카 남서부 연안에 서식하는 가장 작은 해양 포유류 중 하나로, 갑각류, 연체동물, 어류 등을 사냥하며 과거 남획으로 개체 수가 급감하여 현재는 보호받고 있지만 서식지 파괴 등의 위협에 직면해 있다. - 고래 - 등지느러미
등지느러미는 수생 척추동물의 등 중앙선에 위치하여 몸의 좌우 기울임을 방지하고 방향 전환을 돕는 짝을 이루지 않는 지느러미이며, 일부 어종은 추진력, 먹이 유인 등 다양한 용도로 사용하고 종을 식별하는 데 활용되기도 한다. - 고래 - 고래의 노래
고래의 노래는 고래가 내는 다양한 소리를 의미하며, 이빨고래류는 반향정위와 의사소통을 위해 고주파 소리를, 수염고래류는 성적 선택과 관련된 저주파 소리를 내며, 해양 소음 공해의 영향을 받는다.

2. 진화 및 분류
고래는 몸 크기에 따라 크게 긴수염고래류와 이빨고래류로 나뉜다. 몸길이는 작은 것은 1.5m 정도이나 큰 것은 31m에 이른다.[179]
분자생물학, 면역학 및 화석 연구에 따르면 고래목(Cetacea)은 발굽이 두 개인 우제류와 계통적으로 가까운 관계이며, 하마가 가장 가까운 현존 친척이다.[76] 고래의 직계 계통은 약 5,580만 년 전 초기 에오세(Eocene)에 초기 우제류와 함께 시작되었다.[76]
2009년 스폴딩(Spaulding) 등의 연구에 기초한 계통 분류는 다음과 같다.[179]
{| class="wikitable"
|-
! 우제목/경우제목
|-
|
{| class="wikitable"
|-
|낙타과
|-
! 전찬류
|-
|
{| class="wikitable"
|-
! 돼지아목
|-
|

|-
! 경반추류
|-
|
{| class="wikitable"
|-
! 반추류
|-
|
{| class="wikitable"
|-
| 작은사슴과
|-
! 진반추류
|-
|
{| class="wikitable"
|-
| 가지뿔영양과
|-
| 기린과
|-
|
{| class="wikitable"
|-
| 사슴과
|-
|


|}
|}
|}
|-
! 경하마형류/경요치류
|-
|




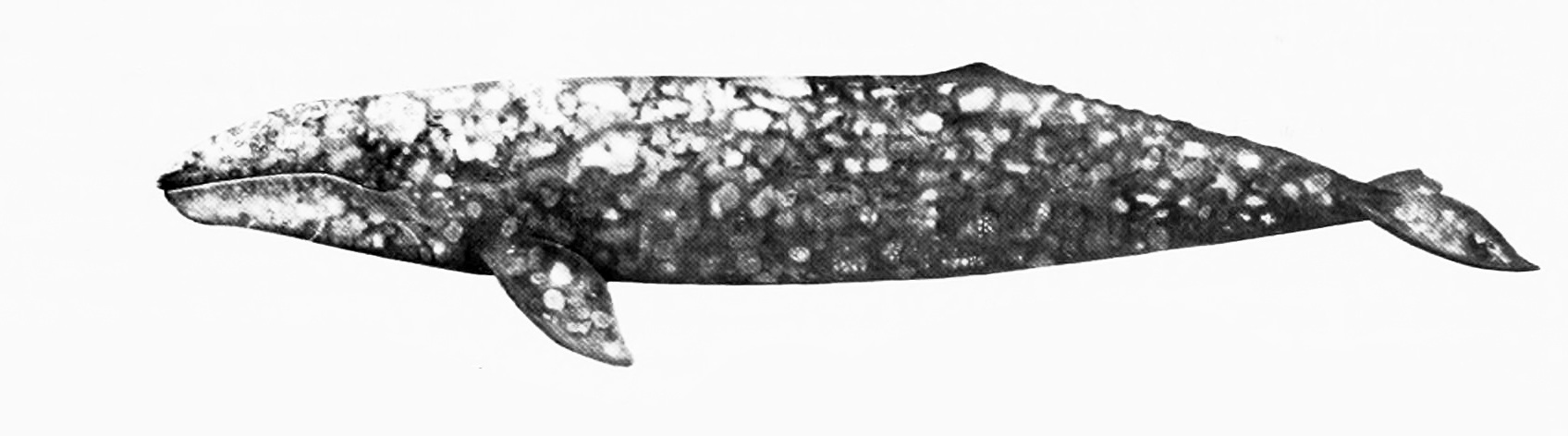






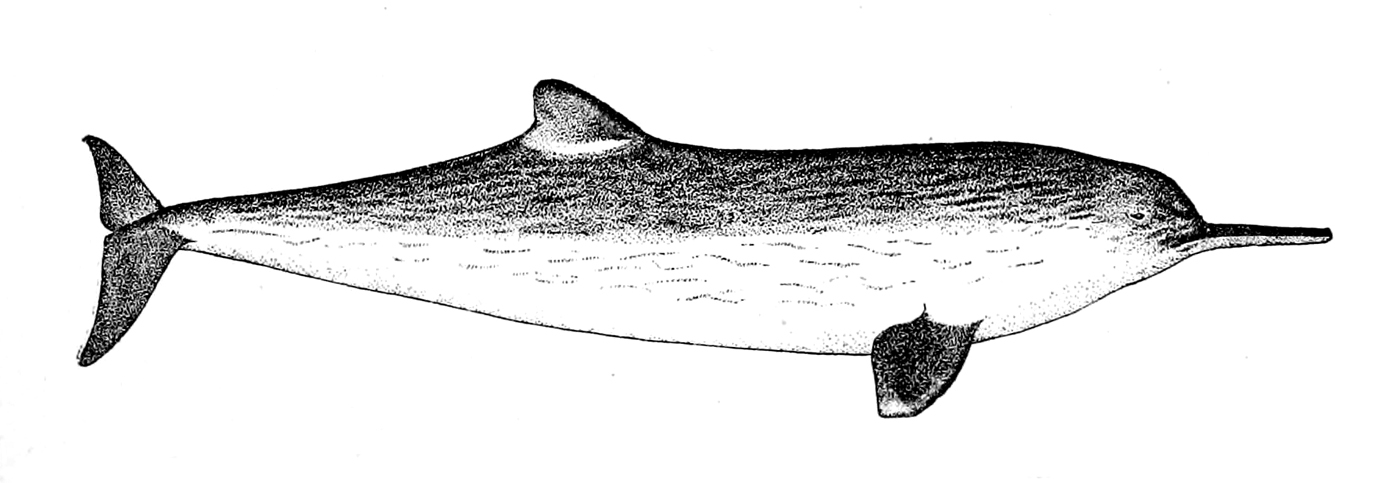



'고래'와 '돌고래'는 비공식적인 용어이다. 한국어에서는 몸길이 5m 이하의 작은 이빨고래류 일부를 돌고래라고 부르지만, 생물 분류학적으로 고래와 돌고래 사이의 명확한 경계는 없다.[135]
2. 1. 진화
과학자들은 고래가 육지 동물이었을 것으로 추정하는데, 그 근거로 폐호흡과 육상 동물의 손가락뼈 5개를 가지고 있다는 점을 든다. 고래의 가슴지느러미에서 손가락뼈를 찾아볼 수 있으며, 콧구멍은 고래가 바다 속에서 계속 생활하기 위해 주기적으로 수면으로 올라와 공기 호흡을 하는 생리 현상의 기능을 한다.[182][183] 고래는 수면에서 60분을 기준으로 약 10분 전후로 호흡하며, 수중에서 약 50분가량 머무를 수 있는 것으로 알려져 있다.수염고래(Mysticeti)와 이빨고래(Odontoceti)의 두 아목은 약 3,400만 년 전에 갈라진 것으로 여겨진다.[13]
수염고래는 이빨 대신 케라틴으로 만들어진 털을 가지고 있다. 이 털은 바닷물에서 크릴과 다른 작은 무척추동물을 걸러낸다. 귀신고래는 저서성 연체동물을 먹는다. 참고래과(balaenopterids)는 목주름을 이용하여 입을 벌려 먹이를 섭취하고 물을 걸러낸다. 긴수염고래과(북극고래와 대왕고래)는 머리가 거대하여 체중의 40%를 차지할 수 있다. 대부분의 수염고래는 북반구와 남반구의 먹이가 풍부한 차가운 바다를 선호하며, 출산을 위해 적도로 이동한다. 이 과정에서 지방 저장량에 의존하여 몇 달 동안 단식할 수 있다.
이빨고래 아목(Odontocetes)에는 향유고래, 부리고래, 범고래, 돌고래, 쇠돌고래 등이 포함된다. 일반적으로 이들의 이빨은 물고기, 오징어 또는 다른 해양 무척추동물을 잡는 데 진화했지만 씹는 데는 사용되지 않으므로 먹이는 통째로 삼킨다. 이빨의 모양은 원뿔형(돌고래와 향유고래), 삽 모양(쇠돌고래), 못 모양(흰돌고래), 엄니 모양(일각돌고래) 또는 다양한 모양(수컷 부리고래)을 한다. 암컷 부리고래의 이빨은 잇몸에 숨겨져 보이지 않으며, 수컷 부리고래 대부분은 짧은 엄니 두 개만 가지고 있다. 일각돌고래는 엄니 외에는 흔적 이빨을 가지고 있으며, 이 엄니는 수컷과 암컷의 15%에 존재하며 수온, 압력, 염도를 감지하는 수백만 개의 신경을 가지고 있다. 범고래와 같이 일부 이빨고래는 바다표범류와 다른 고래와 같은 포유류를 먹는다.
이빨고래는 발달된 감각을 가지고 있다. 시력과 청력은 공기와 물 모두에 적응되어 있으며, 멜론을 사용하여 고도의 소나 능력을 가지고 있다. 청력은 공기와 물 모두에 매우 잘 적응되어 있어 일부 눈이 먼 개체도 생존할 수 있다. 향유고래와 같은 일부 종은 깊은 곳까지 잠수하는 데 잘 적응되어 있다. 이빨고래의 여러 종은 수컷과 암컷이 다른 성적 이형을 보이는데, 이는 일반적으로 성적 과시 또는 공격을 위한 것이다.
오늘날 고래류의 직계 조상은 아마도 가장 유명한 종인 *Dorudon*을 포함하는 Dorudontidae에서 찾을 수 있을 것이다. *Dorudon*은 *Basilosaurus*와 같은 시대에 살았다. 두 그룹 모두 이미 포유류의 고막을 대체하는 고정된 이소골(bulla)과 수중 방향성 청력을 위한 소리 전달 요소와 같이 오늘날 고래의 전형적인 해부학적 특징을 일부 발달시켰다. 그들의 손목은 단단해졌고, 아마도 지느러미의 전형적인 구조에 기여했을 것이다. 그러나 뒷다리는 존재했지만 크기가 상당히 줄어들었고 흔적 기관화된 골반과 연결되어 있었다.[76]
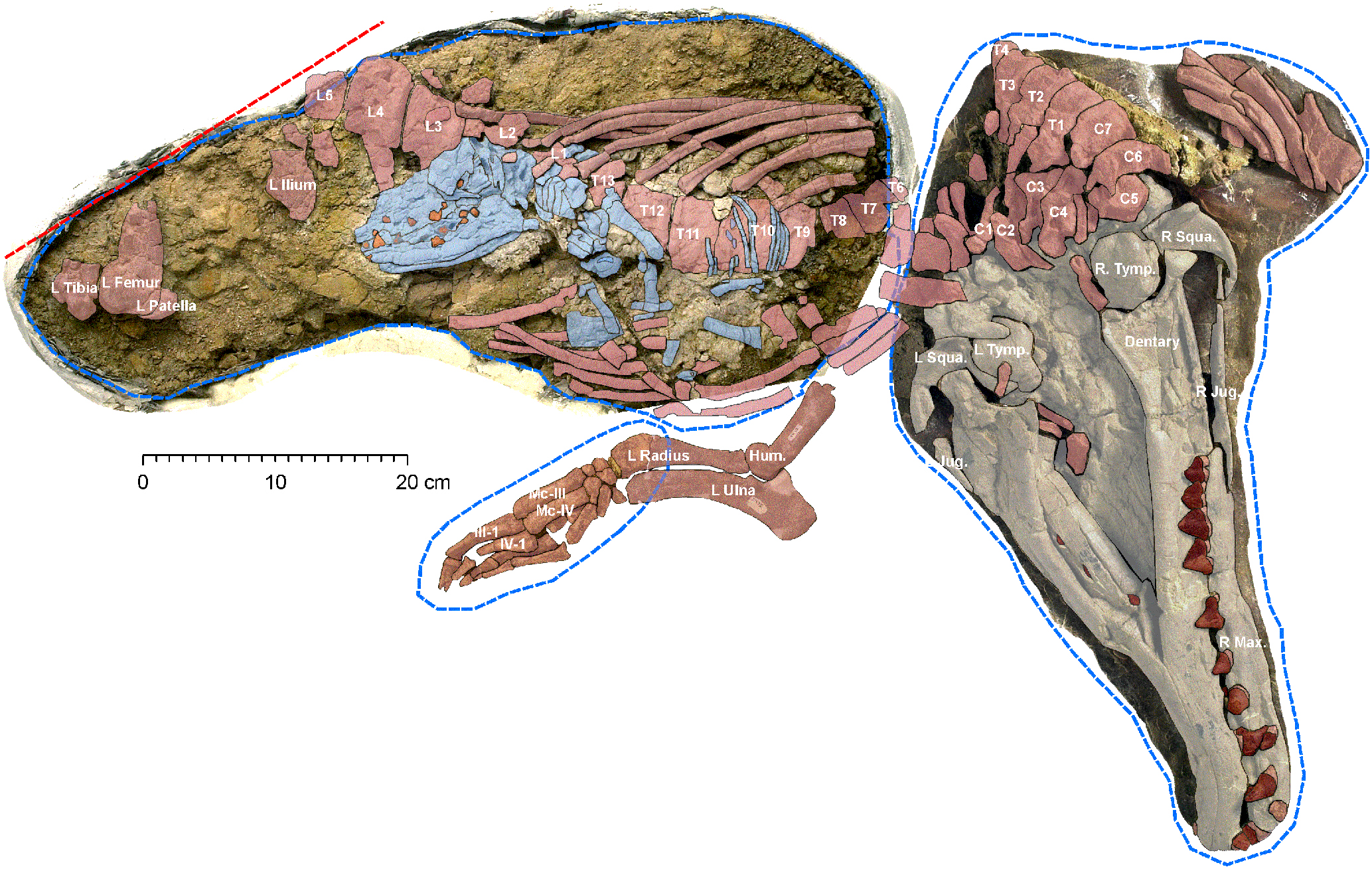
화석 기록은 육지에서 수중 생활로의 점진적인 이행 과정을 보여준다. 뒷다리가 퇴화하면서 척추의 유연성이 증가했다. 이로 인해 고래는 꼬리가 수직으로 물을 치면서 이동할 수 있게 되었다. 앞다리는 지느러미로 변형되었고, 육지에서의 이동성은 상실되었다.
가장 오래된 고래류(고래상목) 중 하나는 파키스탄 중신세 지층에서 발견된 *파키케투스*이다. 늑대 크기의 동물이며, 골격은 부분적으로만 알려져 있다. 기능적인 다리를 가지고 있었고 해안 근처에 서식했는데, 이는 여전히 육지에서 이동할 수 있었음을 시사한다. 긴 주둥이는 육식성 치아를 가지고 있었다.[76]
육지에서 바다로의 이행은 약 4,900만 년 전으로 거슬러 올라가는데, 파키스탄에서 발견된 "걷는 고래"인 *암불로케투스*가 그 예이다. 길이는 최대 3m에 달했다. 이 고대 고래의 다리는 다리 모양이었지만, 이미 완전히 수중 생활을 했는데, 이는 육지 생활과 무관한 생활 방식으로의 전환이 매우 빠르게 일어났음을 나타낸다.[73] 주둥이는 길고, 콧구멍과 눈은 머리 위쪽에 있었다. 꼬리는 강력했고, 물속에서의 이동을 지원했다. *암불로케투스*는 아마도 기수역의 맹그로브 숲에 살았고, 하안지역에서 어류와 다른 척추동물을 포식하는 포식자였을 것이다.[74]
약 4,500만 년 전의 종으로는 *인도케투스*, *쿠치케투스*, *로도케투스*, *앤드류시피우스* 등이 있는데, 모두 물에서의 생활에 적응한 종들이다. 이 종들의 뒷다리는 퇴화했고, 몸의 형태는 현대 고래와 유사하다. 프로토케투스과에 속하는 *로도케투스*는 완전히 수중 생활을 한 최초의 종으로 여겨진다. 몸은 유선형이고 가늘었으며, 손과 발의 뼈가 길었다. 융합된 골반 요추가 있어 꼬리의 부유 운동을 지지할 수 있었다. 아마도 훌륭한 수영 선수였지만, 현대의 물개처럼 육지에서는 서투르게 움직였을 가능성이 크다.[76]
약 4천만 년 전, 에오세 후기부터 고래류는 아열대 해양에 서식하며 더 이상 육지로 올라오지 않았다. 예를 들어, 길이 18미터에 달하는 바실로사우루스(Basilosaurus)가 있는데, 이는 때때로 ''제우글로돈''(Zeuglodon)이라고도 불린다. 육지에서 바다로의 이행은 약 1천만 년 만에 완료되었다. 이집트의 와디 알 히탄("고래 계곡")에는 ''바실로사우루스''의 골격을 비롯한 다른 해양 척추동물의 골격이 다수 발견된다.[75]
분자생물학, 면역학, 그리고 화석 연구는 고래목(Cetacea)이 발굽이 두 개인 우제류(Artiodactyla)와 계통적으로 가까운 관계임을 보여준다. 고래의 직계 계통은 약 5,580만 년 전 초기 에오세(Eocene)에 초기 우제류와 함께 시작되었다.[76] 대부분의 분자생물학적 증거는 하마과(Hippopotamidae)의 하마가 가장 가까운 현존하는 친척임을 시사한다. 공통적인 해부학적 특징으로는 후구치(molars)의 형태학적 유사성, 측두골(temporal bone)의 골륜(bulla), 그리고 이전에는 고래목에서만 발견되는 것으로 알려졌던 두개골 특징인 외피(involucre) 등이 있다.[76] 화석 기록에 따르면 형태학적으로 구별되는 하마 계통은 약 1,500만 년 전에 나타났기 때문에, 고래목과 하마는 형태학적으로 서로 다른 공통 조상으로부터 갈라져 나왔을 것이다.[77][78][79] 가장 두드러진 공통적인 특징은 발목 윗부분의 뼈인 거골(talus bone)이다. 초기 고래류인 고래하목(archaeocetes)은 짝수 발가락을 가진 우제류에서만 발견되는 이중 거골(double castors)을 보여준다. 이와 같은 발견은 인도와 파키스탄 북부의 테티스해(Tethys Sea) 퇴적물에서 나왔다. 테티스해는 아시아 대륙과 북쪽으로 이동하는 인도판 사이의 얕은 바다였다.

분자 및 형태학적 증거는 전통적으로 정의된 우제류(artiodactyl)가 고래목에 대해서 단계통군(paraphyletic)임을 시사한다. 고래목은 우제류 내부에 깊이 자리 잡고 있으며, 두 그룹은 공통 조상을 가진 자연 분류군인 분지군(clade)을 형성하는데, 이를 위해 때때로 Cetartiodactyla라는 이름이 사용된다. 현대 분류법은 우제류(또는 고래우제류)를 네 개의 하위 분류군으로 나눈다. 낙타류(Tylopoda), 돼지와 페카리(Suina), 반추동물(Ruminantia), 그리고 하마와 고래(Whippomorpha)이다. 고래목이 우제류 내에 위치한다는 추정은 다음과 같은 분지도(cladogram)에서 나타낼 수 있다.[80][81][82][83][84]
{| class="wikitable"
|-
! 우제류
|-
|
{| class="wikitable"
|-
| 낙타류 (낙타)
|-
! Artiofabula
|-
|
{| class="wikitable"
|-
| 돼지과 (돼지)
|-
! Cetruminantia
|-
|
{| class="wikitable"
|-
! 반추동물 (반추동물)
|-
|

|-
! Cetancodonta/Whippomorpha
54 mya
|-
|

|}
|}
|}
|}
다음은 2009년 스폴딩(Spaulding) 등의 연구에 기초한 계통 분류이다.[179]
{| class="wikitable"
|-
! 우제목/경우제목
|-
|
{| class="wikitable"
|-
|낙타과
|-
! 전찬류
|-
|
{| class="wikitable"
|-
! 돼지아목
|-
|
|-
! 경반추류
|-
|
{| class="wikitable"
|-
! 반추류
|-
|
{| class="wikitable"
|-
| 작은사슴과
|-
! 진반추류
|-
|
{| class="wikitable"
|-
| 가지뿔영양과
|-
| 기린과
|-
|
{| class="wikitable"
|-
| 사슴과
|-
|
|}
|}
|}
|-
! 경하마형류/경요치류
|-
|
|}
|}
|}
|}
2. 2. 분류
고래는 크게 긴수염고래류와 이빨고래류로 나뉜다.긴수염고래류는 태생기에 이가 나지만, 출생하기 전 퇴화·흡수되어 평생 이빨이 나지 않는다. 대신 입천장에 난 고래수염이 먹이를 걸러내는 소쿠리 구실을 한다. 두개골은 좌우 대칭이며, 외비공(外鼻孔)은 두 개이다. 대부분 일부일처제이며 크게 무리를 짓지 않는다. 플랑크톤성 작은 갑각류를 즐겨 먹는다. 임신기간은 10~12개월이며, 새끼는 성숙한 몸길이의 1/3 크기로 태어난다. 번식은 2년에 1회 하며 성적으로 성숙하는 데 8~10년이 걸리고, 수명은 80년 전후로 추정된다. 긴수염고래류는 포경업의 포획 대상이었으나, 현재는 포획 금지로 보호받고 있다.[179]
이빨고래류는 이빨로 물고기, 오징어 등을 잡지만 씹지 않고 통째로 삼킨다. 이빨 모양은 돌고래와 향유고래의 원뿔형, 쇠돌고래의 삽 모양, 흰돌고래의 못 모양, 일각돌고래의 엄니 모양 등 다양하다. 이빨고래는 발달된 감각, 특히 멜론을 이용한 고도의 소나 능력을 가지고 있다. 일부 종은 깊은 잠수에 적응되어 있으며, 수컷과 암컷의 형태가 다른 성적 이형을 보인다.
수염고래와 이빨고래는 약 3,400만 년 전에 갈라졌다.[13] 분자생물학, 면역학, 화석 연구에 따르면 고래목은 우제류와 가까운 관계이며, 하마가 가장 가까운 현존 친척이다.[76] 고래의 직계 계통은 약 5,580만 년 전 초기 에오세(Eocene)에 초기 우제류와 함께 시작되었다.[76]
분류[85]
고래의 몸은 유선형으로 헤엄치기에 적합한 형태이다. 목은 굵고 짧아, 큰 종류는 목을 돌리는 동작조차 어렵다. 꼬리부분도 굵고 커서 몸통 부분과의 경계를 찾기 어렵다. 꼬리 뒤 끝에는 수평으로 뻗은 꼬리지느러미가 있고, 꼬리의 상하운동으로 주된 유영력을 일으킨다. 앞다리는 가슴지느러미로 변했는데, 긴수염고래류의 일부를 제외하고는 가슴지느러미 속에 다섯 발가락의 골격이 들어 있다. 뒷다리는 퇴화해서 몸 밖으로 나와 있지 않으며, 등의 살갗이 불룩하게 솟아서 생긴 등지느러미를 가진 종류가 많다. 귓바퀴는 없으며, 외계와 고막으로 접하지 않으므로 귓구멍은 살갗 밑에서 끊어져 있으나, 음파는 몸의 모든 부분으로부터 직접 속귀로 전달되며 음감각이 뛰어나다. 눈은 몸의 양 측면에 있다.[178]
'고래'와 '돌고래'는 비공식적인 용어이다. 한국어에서는 몸길이 5m 이하의 작은 이빨고래류 일부를 돌고래라고 부르지만, 생물 분류학적으로 고래와 돌고래 사이의 명확한 경계는 없다.[135]
3. 해부학적 특징

고래는 외형은 어류에 가깝지만, 새끼고래는 자궁 안에서 자라고 태어난 뒤에는 어미고래의 젖을 먹고 자란다. 폐로 호흡하고 온혈동물이며, 체온은 생체근육 속에서 36-38°C로 일정하다.
고래의 피부는 매끄럽고 탄력이 있어 물 속을 쉽게 미끄러져 나아간다. 또한 피부 밑에 지방층이 있어서 체온을 따뜻하게 유지할 수 있다. 예를 들어 흑고래는 지방의 두께가 50cm나 되어, 먹이가 부족할 때는 지방을 분해하여 오랫동안 견딜 수 있다. 지방은 물보다 가벼워 고래를 떠오르게 하는 부력도 증가시킨다.[178]
고래는 규칙적으로 물 표면으로 올라와 숨을 쉬어야 한다. 수염고래는 대개 5-15분마다 숨을 쉬지만, 40분 동안 숨을 쉬지 않고도 견딜 수 있다. 향고래는 2시간까지 숨을 안 쉴 수도 있다. 고래가 오랫동안 숨을 쉬지 않고도 견딜 수 있는 것은 고래의 근육이 다른 포유류의 근육보다 산소를 많이 저장하기 때문이다. 물 밖으로 나왔을 때는 다시 잠수하기 전에 몇 차례의 숨쉬기로 조직 내에 산소를 가득 채워야 한다.[178]
고래는 물 표면으로 올라와서 물보라를 일으키며 앞으로 구른다. 이 2초 정도의 시간에 2000L의 공기를 들이마신다. 고래는 머리꼭대기에 있는 숨구멍인 분기공으로 숨을 쉰다. 이빨고래는 분수공이 하나이고, 수염고래는 둘이다. 고래가 숨을 쉴 때는 강력한 근육과 판에 의해 분수공이 넓게 열리지만, 숨을 쉰 후에는 다시 단단히 닫힌다. 고래가 숨을 내쉴 때, 분수처럼 물보라가 일어난다. 분수 기둥은 주로 수증기로 이루어지지만, 점액질과 기름 방울도 섞여 있다.[178]
과학자들은 고래가 육지동물이었을 것으로 여기는데, 그 이유는 폐호흡과 육상동물이 가진 5개의 손가락뼈를 보유하고 있기 때문이다. 손가락뼈는 고래의 가슴지느러미에서 찾아볼 수 있으며, 폐호흡을 하는 콧구멍은 고래가 바다 속에서 계속 생활하기 위해 주기적으로 수면으로 올라왔을 때 공기호흡을 하는 생리현상의 기능을 한다.[182][183]
고래류는 유선형의 몸을 가지고 있으며, 앞다리는 지느러미이다. 거의 모든 종이 등에 등지느러미를 가지고 있지만, 종에 따라 다양한 형태를 취할 수 있다. 흰돌고래와 같이 등지느러미가 없는 몇몇 종도 있다. 지느러미와 등지느러미 모두 물속에서 안정성과 조향을 위해 사용된다.
수컷의 생식기와 암컷의 유선은 몸속으로 들어가 있다.[15][16] 수컷의 생식기는 흔적만 남은 골반에 붙어 있다.[17]
몸은 지방층으로 알려진 두꺼운 지방층으로 덮여 있다. 이것은 단열을 제공하고 고래류에게 매끄럽고 유선형의 몸매를 제공한다. 큰 종에서는 두께가 최대 0.5m에 이를 수 있다.
많은 이빨고래류에서 성적 이형이 진화했다. 향유고래, 일각돌고래, 많은 부리고래과 종들, 몇몇 돌고래과 종들, 범고래, 낫돌고래, 동부 회색돌고래, 북방긴수염돌고래가 이러한 특징을 보인다.[18] 이러한 종의 수컷은 암컷에게 없는 외부 특징을 발달시켰는데, 이는 전투나 과시에 유리하다. 예를 들어, 수컷 향유고래는 암컷보다 최대 63% 더 크며, 많은 부리고래는 수컷 간의 경쟁에 사용되는 엄니를 가지고 있다.[18][19]
고래류에는 뒷다리가 없으며, 귓바퀴나 털과 같은 다른 외부 부속물도 없다.[20]
3. 1. 머리
고래는 특히 수염고래의 경우 넓게 튀어나온 아래턱 때문에 머리가 길다. 북극고래의 수염판은 길이가 9m에 달할 수 있다. 콧구멍은 분출공을 형성하며, 이빨고래는 하나, 수염고래는 두 개이다.[21]
콧구멍은 눈 위쪽 머리 꼭대기에 위치하여 공기를 마시기 위해 수면으로 올라올 때 몸의 나머지 부분은 물속에 잠긴 상태를 유지할 수 있다. 후두골은 상당히 짧아지고 변형되어 있다. 콧구멍이 머리 꼭대기로 이동함에 따라 비강은 두개골을 수직으로 통과한다.[22] 위턱의 이빨이나 수염판은 상악골에만 위치한다.
이빨고래에서는 머리 버클 역할을 하는 멜론에 결합 조직이 존재한다. 이것은 부력과 생체 음파 탐지를 돕는 공기주머니와 지방으로 채워져 있다. 향유고래는 특히 멜론이 두드러지는데, 이것은 향유주머니라고 하며, 동명의 경랍을 포함하고 있어 "향유고래"라는 이름이 붙었다. 일각돌고래의 긴 엄니조차도 변형된 이빨이다.
수염고래의 수염은 긴 섬유질 케라틴 가닥으로 구성되어 있다. 이빨 대신에 위치하며 거대한 술과 같은 모양을 하고 있으며, 물에서 플랑크톤과 크릴을 걸러내는 데 사용된다.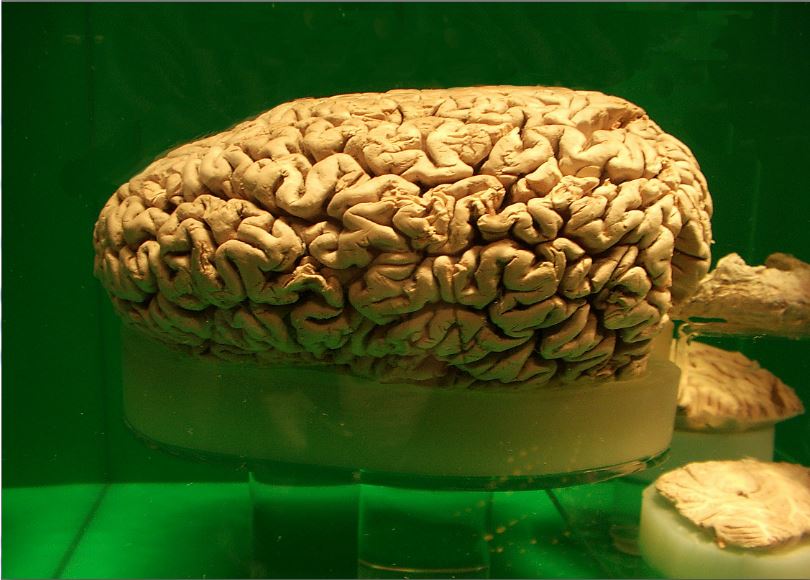
향유고래는 지구상의 어떤 동물보다도 가장 큰 뇌 질량을 가지고 있으며, 성체 수컷의 평균 뇌 크기는 8000cm3이고 무게는 7.8kg이다.[23] 흰돌고래와 일각돌고래와 같은 일부 이빨고래류의 뇌-체중 비율은 인간 다음으로 높다.[24]
뇌 크기는 이전에는 지능의 주요 지표로 간주되었다. 뇌의 대부분은 신체 기능 유지를 위해 사용되기 때문에, 뇌-체중 비율이 높을수록 인지 활동에 사용할 수 있는 뇌 질량이 증가할 수 있다.
3. 2. 몸
고래의 몸은 유선형으로 헤엄치기에 적합한 형태이다. 목은 굵고 짧아, 큰 종류는 목을 돌리는 동작조차 어렵다. 입은 크고, 콧구멍은 머리 꼭대기에 있다. 꼬리부분도 굵고 커서 몸통 부분과의 경계를 찾기 어렵다. 꼬리 뒤 끝에는 수평으로 뻗은 꼬리지느러미가 있고, 꼬리의 상하운동으로 주된 유영력을 일으킨다. 앞다리는 가슴지느러미로 변했는데, 긴수염고래류의 일부를 제외하고는 가슴지느러미 속에 다섯 발가락의 골격이 들어 있다. 뒷다리는 퇴화해서 몸 밖으로 나와 있지 않으며, 등의 살갗이 불룩하게 솟아서 생긴 등지느러미를 가진 종류가 많다. 귓바퀴는 없으며, 외계와 고막으로 접하지 않으므로 귓구멍은 살갗 밑에서 끊어져 있으나, 음파는 몸의 모든 부분으로부터 직접 속귀로 전달되며 음감각이 뛰어나다. 눈은 몸의 양 측면에 있다.[178]
고래는 외형은 어류에 가깝지만, 새끼고래는 자궁 안에서 자라고 태어난 뒤에는 어미고래의 젖을 먹고 자란다. 폐로 호흡하고 온혈동물이며, 체온은 생체근육 속에서 36-38 °C로 일정하다.
고래의 피부는 매끄럽고 탄력이 있어 물 속을 쉽게 미끄러져 나아간다. 또한 피부 밑에 지방층이 있어서 체온을 따뜻하게 유지할 수 있다. 예를 들어 흑고래는 지방의 두께가 50cm나 되어, 먹이가 부족할 때는 지방을 분해하여 오랫동안 견딜 수 있다. 지방은 물보다 가벼워 고래를 떠오르게 하는 부력도 증가시킨다.[178]
고래는 폐로 호흡하기 때문에 규칙적으로 물 표면으로 올라와 숨을 쉬어야 한다. 수염고래는 대개 5-15분마다 숨을 쉬지만, 40분 동안 숨을 쉬지 않고도 견딜 수 있다. 향고래는 2시간까지 숨을 안 쉴 수도 있다. 고래가 오랫동안 숨을 쉬지 않고도 견딜 수 있는 것은 고래의 근육이 다른 포유류의 근육보다 산소를 많이 저장하기 때문이다. 물 밖으로 나왔을 때는 다시 잠수하기 전에 몇 차례의 숨쉬기로 조직 내에 산소를 가득 채워야 한다.[178]
고래는 숨을 쉬기 위해 물 표면으로 올라와서 물보라를 일으키며 앞으로 구른다. 이 2초 정도의 시간에 2000L의 공기를 들이마신다. 고래는 머리꼭대기에 있는 숨구멍인 분기공으로 숨을 쉰다. 이빨고래는 분수공이 하나이고, 수염고래는 둘이다. 고래가 숨을 쉴 때는 강력한 근육과 판에 의해 분수공이 넓게 열리지만, 숨을 쉰 후에는 다시 단단히 닫힌다. 고래가 숨을 내쉴 때, 분수처럼 물보라가 일어난다. 분수 기둥은 주로 수증기로 이루어지지만, 점액질과 기름 방울도 섞여 있다.[178]
과학자들은 고래가 육지동물이었을 것으로 여기는데, 그 이유는 폐호흡과 육상동물이 가진 5개의 손가락뼈를 보유하고 있기 때문이다. 손가락뼈는 고래의 가슴지느러미에서 찾아볼 수 있으며, 폐호흡을 하는 콧구멍은 고래가 바다 속에서 계속 생활하기 위해 주기적으로 수면으로 올라왔을 때 공기호흡을 하는 생리현상의 기능을 한다.[182][183] 고래의 수면에서의 호흡은 60분을 기준으로 약 10분 전후이며 고래는 수중에서 약 50분가량 머무를 수 있는 것으로 알려져 있다.
고래류의 몸은 일반적으로 물고기와 비슷하며, 이는 그들의 생활 방식과 서식 환경에 기인한다. 다른 고등 포유류(진수하강)와 중요한 특징을 공유하지만, 그들의 몸은 서식지에 잘 적응되어 있다.[14]
고래류는 유선형의 몸을 가지고 있으며, 앞다리는 지느러미이다. 거의 모든 종이 등에 등지느러미를 가지고 있지만, 종에 따라 다양한 형태를 취할 수 있다. 흰돌고래와 같이 등지느러미가 없는 몇몇 종도 있다. 지느러미와 등지느러미 모두 물속에서 안정성과 조향을 위해 사용된다.
수컷의 생식기와 암컷의 유선은 몸속으로 들어가 있다.[15][16] 수컷의 생식기는 흔적만 남은 골반에 붙어 있다.[17]
몸은 지방층으로 알려진 두꺼운 지방층으로 덮여 있다. 이것은 단열을 제공하고 고래류에게 매끄럽고 유선형의 몸매를 제공한다. 큰 종에서는 두께가 최대 1m에 이를 수 있다.
많은 이빨고래류에서 성적 이형이 진화했다. 향유고래, 일각돌고래, 많은 부리고래과 종들, 몇몇 돌고래과 종들, 범고래, 낫돌고래, 동부 회색돌고래, 북방긴수염돌고래가 이러한 특징을 보인다.[18] 이러한 종의 수컷은 암컷에게 없는 외부 특징을 발달시켰는데, 이는 전투나 과시에 유리하다. 예를 들어, 수컷 향유고래는 암컷보다 최대 63% 더 크며, 많은 부리고래는 수컷 간의 경쟁에 사용되는 엄니를 가지고 있다.[18][19]
고래류에는 뒷다리가 없으며, 귓바퀴나 털과 같은 다른 외부 부속물도 없다.[20]
고래류는 모두 수생(水生)이며 주로 바다에 서식하지만, 강돌고래류 등 일부는 강이나 기수역에 서식한다.[144] 현생 동물 중에서는 몸길이와 몸무게가 가장 큰 그룹을 포함하며, 특히 흰긴수염고래(Balaenoptera musculus)는 동물로서 역사상 가장 큰 질량(몸무게 약 130ton)을 자랑한다. 반면, 고래목 중에서 가장 작은 것은 코가시라네ズミ돌고래(Phocoena phocoena)이며, 몸길이는 약 1.5m, 몸무게는 50kg 정도에 불과하다. 어류나 오징어류 등의 두족류를 먹는 이빨고래류와, 크릴 등의 플랑크톤이나 무리를 지어 사는 작은 물고기를 먹는 수염고래류는 먹이 습성이 다르지만, 모두 넓은 의미에서 육식성이다. 이빨고래류는 해양 생태계의 최상위를 비롯해 상위 포식자의 많은 부분을 차지하며, 수염고래류도 하위 소비자의 최대 종을 포함하는 거대한 그룹으로서 다양한 진화에 성공했다.
고래류의 귀에는 귓바퀴가 없고, 단지 직경 2mm 정도의 구멍일 뿐이며, 귀지가 막혀 있다. 수염고래의 경우 귀지의 층을 세어 어느 정도의 나이를 추정할 수 있다.[145] 이에 반해 이빨고래의 나이 추정에는 이빨이 사용된다.[145] 지방을 축적하여 수분을 만들어 생활한다. 땀샘은 없다. 머리의 등쪽에 호흡을 위한 분기공이 있다. 분기공은 수염고래에서는 2개, 이빨고래에서는 1개이다. 분기공은 개폐가 가능하며, 머리를 수면 위로 내밀고 분기공을 열어 공기를 들이마시고, 그 외 잠수할 때는 분기공을 닫아 물이 들어가는 것을 막는다.[146] 코를 골기도 한다. 똥은 고형분이 적고 액체에 가깝다. 포유류이므로 항온동물이며, 체온은 35~36℃로 일정하게 유지된다.[147] 배꼽도 있다. 젖꼭지는 2개이며, 감기에 걸리기도 한다. 수영 속도는 시속 3km에서 50km 정도이다. 지금까지 알려진 최고령은 대왕고래가 114세, 흰긴수염고래가 110세이며, 소형종이 될수록 최고령은 낮아지는 경향이 있다.[148]
3. 3. 호흡
고래는 폐로 호흡하기 때문에 규칙적으로 물 표면으로 올라와 숨을 쉬어야 한다. 수염고래는 대개 5-15분마다 숨을 쉬지만, 40분 동안 숨을 쉬지 않고도 견딜 수 있다. 향고래는 2시간까지 숨을 안 쉴 수도 있다.[178] 고래가 오랫동안 숨을 쉬지 않고도 견딜 수 있는 것은 고래의 근육이 다른 포유류의 근육보다 산소를 많이 저장하기 때문이다. 물 밖으로 나왔을 때는 다시 잠수하기 전에 몇 차례의 숨쉬기로 조직 내에 산소를 가득 채워야 한다.
고래는 숨을 쉬기 위해 물 표면으로 올라와서 물보라를 일으키며 앞으로 구른다. 이 2초 정도의 시간에 2,000L의 공기를 들이마신다. 고래는 머리꼭대기에 있는 숨구멍인 분수공으로 숨을 쉰다. 이빨고래는 분수공이 하나이고, 수염고래는 둘이다. 고래가 숨을 쉴 때는 강력한 근육과 판에 의해 분수공이 넓게 열리지만, 숨을 쉰 후에는 다시 단단히 닫힌다. 고래가 숨을 내쉴 때, 분수처럼 물보라가 일어난다. 분수 기둥은 주로 수증기로 이루어지지만, 점액질과 기름 방울도 섞여 있다.[178]
과학자들은 고래가 육지동물이었을 것으로 여기는데, 그 이유로 폐호흡과 육상동물의 손가락뼈 5개를 보유하고 있다는 점을 든다. 손가락뼈는 고래의 가슴지느러미에서 찾아볼 수 있으며, 폐호흡을 하는 콧구멍은 고래가 바다 속에서 계속 생활하기 위해 주기적으로 수면으로 올라왔을 때 공기 호흡을 하는 생리 현상의 기능을 한다.[182][183] 고래의 수면에서의 호흡은 60분을 기준으로 약 10분 전후이며 고래는 수중에서 약 50분가량 머무를 수 있는 것으로 알려져 있다.
3. 4. 순환
고래류의 호흡계와 순환계 구조는 해양 포유류의 생명에 특히 중요하다. 산소 균형이 효율적인데, 한 번의 호흡으로 폐 용량의 최대 90%를 교체할 수 있다. 반면 육지 포유류의 경우 이 값은 일반적으로 약 15%이다. 흡입하는 동안 폐 조직에 흡수되는 산소량은 육지 포유류의 약 두 배이다. 모든 포유류와 마찬가지로 산소는 혈액과 폐에 저장되지만, 고래류에서는 주로 근육을 포함한 다양한 조직에도 저장된다. 근육 색소인 미오글로빈이 효과적인 결합을 제공한다. 이러한 추가적인 산소 저장은 심해 잠수에 필수적이다. 100m 정도의 깊이를 넘어서면 폐 조직은 수압에 의해 거의 완전히 압축되기 때문이다.
3. 5. 감각
고래는 귓바퀴가 없으며, 외계와 고막으로 접하지 않아 귓구멍은 살갗 밑에서 끊어져 있다. 하지만 음파는 몸의 모든 부분으로부터 직접 속귀로 전달되며 음감각이 뛰어나다.[178] 고래류의 눈은 머리 앞쪽이 아닌 옆쪽에 위치한다. 이는 돌고래와 같이 뾰족한 '부리'를 가진 종만이 앞쪽과 아래쪽으로 좋은 양안시를 가진다는 것을 의미한다. 눈물샘에서는 기름진 눈물을 분비하여 눈을 물의 염분으로부터 보호한다. 수정체는 거의 구형이며, 이는 수심 깊은 곳까지 도달하는 최소한의 빛을 초점에 맞추는 데 가장 효율적이다. 이빨고래류는 미각이나 후각 능력이 거의 없거나 전혀 없지만, 수염고래류는 퇴화되었지만 기능하는 후각계 때문에 후각 능력이 있는 것으로 여겨진다.[40] 고래류는 뛰어난 청력을 가지고 있는 것으로 알려져 있다.[41]
아마존강돌고래는 먹이를 감지하기 위해 전기수용을 사용할 수 있다.[42]
외이(外耳)는 귓바퀴(외형적으로 보이는 부분)를 잃었지만, 여전히 좁은 귓구멍을 유지하고 있다. 소리를 전달하는 세 개의 작은 뼈, 즉 이소골은 치밀하고 단단하며, 육상 포유류의 것과는 모양이 다르다. 반고리관은 다른 포유류에 비해 몸집에 대한 크기가 훨씬 작다.[43]
중이와 내이의 뼈 구조인 고실은 두 개의 치밀하고 단단한 뼈(내이골 및 고실골)로 구성되어 있다. 이것은 중이 안의 공간에 위치하며, 이빨고래류(향유고래과를 제외하고)에서는 이 공간이 짙은 거품으로 채워져 있으며, 고실을 완전히 둘러싸고 있고, 인대에 의해서만 두개골에 연결되어 있다. 이것은 귀를 두개골의 뼈를 통해 전달되는 소리로부터 차단할 수 있는데, 이것은 박쥐에서도 발생하는 현상이다.[44]
고래류는 신음 소리, 울음 소리, 휘파람 소리, 딸깍 소리 또는 혹등고래의 '노래'를 사용하여 의사소통을 위해 소리를 사용한다.[42]
이빨고래류(Odontoceti)는 일반적으로 반향정위 능력이 있다.[45] 이들은 물체의 크기, 모양, 표면 특징, 거리 및 움직임을 식별할 수 있다. 완전한 어둠 속에서도 먹이를 찾고, 추격하고, 잡을 수 있다. 대부분의 이빨고래류는 먹이와 비먹이(예: 사람이나 배)를 구별할 수 있으며, 사육 중인 이빨고래류는 예를 들어 크기나 모양이 다른 공을 구별하도록 훈련될 수 있다. 반향정위 클릭에는 각 동물마다 고유한 특징적인 세부 사항이 포함되어 있으며, 이는 이빨고래류가 자신의 클릭과 다른 클릭을 구별할 수 있음을 시사할 수 있다.[46]
반향정위 능력과 관련된 귀 구조의 차이는 고래류에서 발견되지만, 두개골의 비대칭 또한 반향정위에 사용되는 소리를 생성하는 능력에 영향을 미치는 요인으로 밝혀졌다. 반향정위 능력이 없는 수염고래류(Mysticeti)는 두개골과 얼굴 부위가 일반적으로 대칭적인 반면, 이빨고래류는 반향정위 능력과 관련된 비대칭적인 코와 얼굴을 가지고 있다.[47] 비대칭 정도의 차이는 생성되는 소리의 유형과도 상관관계가 있는 것으로 보인다.[48]
수염고래류는 강화제 없이 매우 얇고 넓은 기저막을 달팽이관에 가지고 있어 저주파에서 초저주파까지의 주파수를 처리하도록 귀가 적응되어 있다.[49]
3. 6. 골격
고래류의 골격은 대부분 치밀골로 이루어져 있어 물속에서 안정성을 유지하는 데 도움을 준다. 일반적인 육상 동물의 치밀뼈인 해면골은 더 가볍고 탄력 있는 물질로 대체된다. 많은 부분에서 뼈 요소는 연골이나 지방으로 대체되어 정수압적 특성을 향상시킨다. 귀와 주둥이에는 고래류에만 있는 고밀도의 뼈 구조가 있는데, 이는 도자기와 비슷하며 다른 뼈보다 소리를 더 잘 전달하여 생체 음파탐지에 도움이 된다.
척추뼈의 수는 종에 따라 40개에서 93개까지 다양하다. 모든 포유류에서 볼 수 있는 경추는 7개의 척추뼈로 구성되지만, 축소되거나 융합되어 있어 수영 중 안정성을 높인다. 지느러미는 9개에서 17개의 척추뼈로 구성된 흉추에 의해 지지된다. 흉골은 연골로 이루어져 있으며, 마지막 2~3쌍의 갈비뼈는 연결되지 않고 체벽에 자유롭게 매달려 있다. 안정적인 요추와 꼬리는 나머지 척추뼈를 포함하며, 미추 아래에는 셰브런 뼈가 있다.
앞다리는 노 모양이며, 팔은 짧고 손가락뼈는 길어 운동을 지원한다. 이들은 연골로 연결되어 있다. 둘째와 셋째 손가락은 손가락 마디가 증식하는 과지골을 보인다. 어깨 관절은 아마존강돌고래를 제외한 모든 고래류에서 유일하게 기능하는 관절이다. 쇄골은 완전히 없다.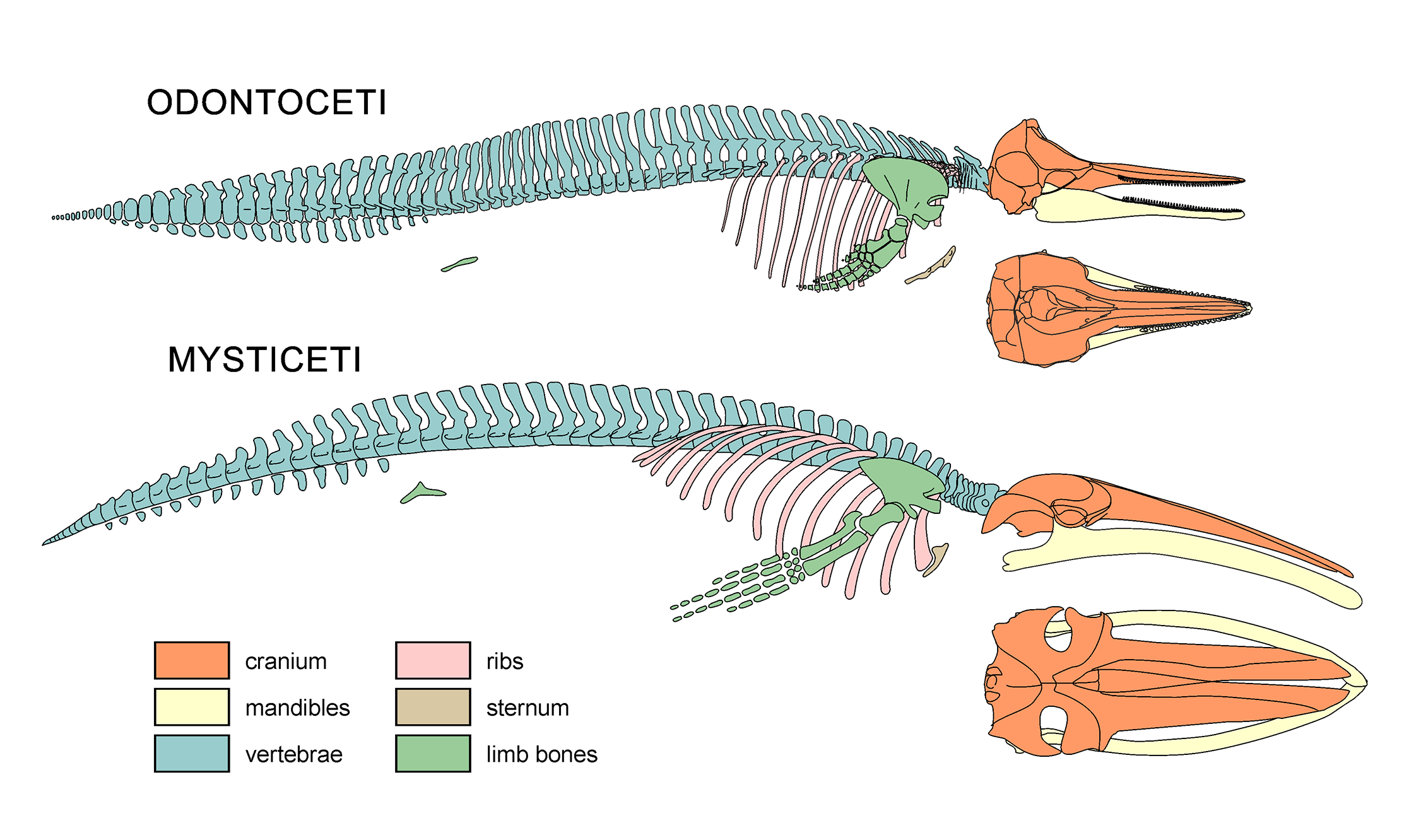
3. 7. 꼬리지느러미

고래류는 추진력을 얻기 위해 꼬리 끝에 연골로 된 꼬리지느러미(fluke)를 가지고 있다. 물고기나 어룡과는 달리 꼬리지느러미는 몸에 수평으로 위치하며 수직 운동으로 사용된다. 어룡과 물고기는 수평으로 움직이는 수직 꼬리를 가지고 있다.[31]
4. 생태
이빨고래류(Odontoceti)는 반향정위 능력이 있어, 물체의 크기, 모양, 특징, 거리, 움직임을 식별하고 어둠 속에서도 먹이를 찾고 추격한다.[45] 대부분의 이빨고래류는 먹이와 비먹이를 구별할 수 있으며, 사육 상태에서는 크기나 모양이 다른 공을 구별하도록 훈련받을 수 있다. 반향정위 클릭에는 각 동물마다 고유한 특징이 있어 자신의 클릭과 다른 클릭을 구별할 수 있다.[46]
수염고래류(Mysticeti)는 달팽이관에 얇고 넓은 기저막을 가지고 있어 저주파에서 초저주파까지의 주파수를 처리하도록 귀가 적응되어 있다.[49] 반면 이빨고래류는 반향정위 능력과 관련된 비대칭적인 코와 얼굴을 가지고 있으며, 비대칭 정도는 생성되는 소리의 유형과 상관관계가 있다.[47][48]
고래류는 잠수하는 동안 심장 활동과 혈액 순환을 낮춰 산소 소비를 줄인다. 긴수염고래는 최대 40분, 향유고래는 60~90분, 낫돌고래는 2시간까지 잠수할 수 있으며, 평균 잠수 깊이는 약 100m이다. 향유고래는 3000m까지 잠수할 수 있지만, 일반적으로는 1200m이다.[56][57]
고래류는 가르치고, 배우고, 협력하고, 계략을 짜고, 슬퍼하는 것으로 알려져 있다.[60] 소형 고래류는 "거품 고리"를 만들거나 물어뜯는 등 복잡한 놀이 행동을 하며, 거품 그물을 만들어 먹이를 모으기도 한다.[61] 대형 고래는 꼬리 지느러미를 물 위로 들어 올리는 "항해"를 하는데, 남방긴수염고래는 아르헨티나와 남아프리카 해안에서, 혹등고래도 이러한 행동을 보인다.
자기 인식은 추상적 사고의 징후로 보이며, 돌고래는 자기 인식을 가지고 있는 것으로 보인다.[63] 거울 테스트를 통해 동물의 자기 인식을 확인하는데, 돌고래는 이 테스트를 통과했다.[64] 그러나 비판적인 시각에서는 클레버 핸스 효과를 주장하며, 옹호론자들은 돌고래의 행동이 다른 개체에 대한 반응과 다르다고 반박한다.[64] 1995년 Marten과 Psarakos는 비디오를 이용해 돌고래의 자기 인식을 테스트하여 사회적 행동이 아닌 자기 인식을 시사한다고 결론지었지만, 이 연구는 복제되지 않았다.[65][64]
고래는 무리를 지어 생활하며, 다양한 소리로 대화한다. 이 소리는 먼 거리에서도 들을 수 있으며, 무리의 개체를 모으는 데 사용된다. 울음소리는 연속된 소리로 20분 정도 지속된 후 반복된다.[178]
의식적으로 숨을 쉬는 고래는 단측구 수면을 하는 것으로 알려져 있는데, 이는 뇌의 한쪽만 자는 것이다. 이를 통해 휴식 중에도 수영, 호흡, 포식자 회피 등을 할 수 있다.[54] 2008년 연구에 따르면, 향유고래는 수직 자세로 얕은 '드리프트 다이빙'을 하며 잠을 자는데, 이때 고래는 접촉하지 않는 한 반응하지 않아 잠수 중 잠을 잘 수 있다는 추측이 나왔다.[55]
집단적인 의사결정은 무리 생활을 하는 여러 고래 종에게 중요하다.[66] 향유고래 무리에서는 복잡한 합의 의사결정이, 범고래와 같은 다른 종에서는 리더십이 존재한다는 증거가 발견되었다.
많은 고래 종들은 계절 서식지를 오가기 위해 회유를 한다. 예를 들어, 귀신고래는 겨울 번식지인 바하칼리포르니아의 따뜻한 석호에서 여름 먹이 지역인 알래스카 연안의 베링해, 추크치해, 보퍼트해까지 약 16000km를 왕복 이동한다.[53]
4. 1. 서식지 및 분포
고래류는 다양한 수생 서식지에서 서식한다. 흰긴수염고래, 혹등고래, 범고래와 같이 거의 모든 바다에 분포하는 종도 있지만, 일부 종은 지역적으로 또는 분포가 끊어진 개체군으로만 나타난다. 바키타는 캘리포니아만의 일부 지역에, 헥터돌고래는 뉴질랜드의 일부 연안 해역에 서식한다. 대부분의 강돌고래 종은 민물에서만 서식한다.[51]브라이드고래나 리스소돌고래처럼 특정 위도, 특히 열대 또는 아열대 해역에 서식하는 종도 많다. 남방긴수염돌고래와 모래시계돌고래는 남극해에서만, 일각돌고래와 흰돌고래는 북극해에서만 서식하는 것처럼 특정 수역에서만 발견되는 종도 있다. 소버비부리고래와 클리메네돌고래는 대서양에만, 태평양흰옆돌고래와 북방긴부리돌고래는 북태평양에만 서식한다.
전 세계적으로 분포하는 종들은 태평양, 대서양, 인도양에서 발견될 수 있다. 그러나 북반구와 남반구 개체군은 시간이 지남에 따라 유전적으로 분리된다. 일부 종에서는 이러한 분리가 결국 남방긴수염고래, 북태평양긴수염고래, 북대서양긴수염고래와 같은 종 분화로 이어진다.[52] 이동성 종의 번식지는 종종 열대 지방에 있으며, 먹이를 찾는 지역은 극지방에 있다.
유럽 해역에서는 이빨고래 25종과 수염고래 7종을 포함하여 32종이 발견된다.
다음은 멸종위기에 처한 야생 동식물종의 국제거래에 관한 협약(워싱턴 조약, CITES) 부속서에 등재된 고래목 종들을 나타낸다.
4. 2. 먹이
태생기에는 이가 나지만, 출생하기까지 퇴화·흡수되고 출생 후에는 평생 이빨이 나지 않는다. 대신 입천장에 난 고래수염이 먹이를 포식하는 데 소쿠리 구실을 한다. 그 때문에 입이 몸의 1/4 내지 1/3이나 된다. 먹이는 떼를 지어 다니는 작은 물고기도 먹지만, 플랑크톤성의 작은 갑각류를 즐겨 먹는다.[181]수염고래는 이빨 대신 케라틴으로 만들어진 털을 가지고 있다. 이 털은 바닷물에서 크릴과 다른 작은 무척추동물을 걸러낸다. 귀신고래는 저서성 연체동물을 먹는다. 참고래과(balaenopterids)는 목주름을 이용하여 입을 벌려 먹이를 섭취하고 물을 걸러낸다. 긴수염고래과(북극고래와 대왕고래)는 머리가 거대하여 체중의 40%를 차지할 수 있다.
이빨고래 아목(Odontocetes)에는 향유고래, 부리고래, 범고래, 돌고래, 쇠돌고래 등이 포함된다. 일반적으로 이들의 이빨은 물고기, 오징어 또는 다른 해양 무척추동물을 잡는 데 진화했지만 씹는 데는 사용되지 않으므로 먹이는 통째로 삼킨다. 이빨의 모양은 원뿔형(돌고래와 향유고래), 삽 모양(쇠돌고래), 못 모양(흰돌고래), 엄니 모양(일각돌고래) 또는 다양한 모양(수컷 부리고래)을 한다. 일부 범고래와 같이 일부 이빨고래는 바다표범류와 다른 고래와 같은 포유류를 먹는다.
무리 생활을 하는 동물들도 사냥을 하는데, 종종 다른 종과 함께 사냥하기도 한다. 많은 돌고래 종들은 큰 참치 무리와 함께 사냥을 하며, 큰 물고기 떼를 따라다닌다. 범고래는 무리를 지어 사냥하며 흰돌고래나 심지어 더 큰 고래를 사냥감으로 삼는다. 혹등고래를 비롯한 다른 고래들은 기포 카펫을 공동으로 만들어 크릴이나 플랑크톤을 떼 지어 몰아넣은 후 덮치기도 한다.[58]
어류나 오징어류 등의 두족류를 먹는 이빨고래류와, 크릴 등의 플랑크톤이나 무리를 지어 사는 작은 물고기를 먹는 수염고래류는 먹이 습성이 다르지만, 모두 넓은 의미에서 육식성이다. 이빨고래류는 해양 생태계의 최상위를 비롯해 상위 포식자의 많은 부분을 차지하며, 수염고래류도 하위 소비자의 최대 종을 포함하는 거대한 그룹으로서 다양한 진화에 성공했다.
4. 3. 번식
많은 종류의 고래는 특정한 계절에만 짝짓기를 한다. 짝짓기 형태는 지느러미발로 서로 치거나 물 위로 뛰어오르거나, 길고 구부러진 지느러미발로 서로 안는 행동을 포함한다. 임신 기간은 보통 10~12개월이며, 한 번에 한 마리의 새끼를 낳지만 드물게 두 마리가 태어나기도 한다. 한두 마리의 다른 암컷이 산모의 출산을 돕기도 한다.[178]고래는 태어날 때부터 몸집이 크다. 갓 태어난 흰긴수염고래는 몸무게가 평균 약 1.8ton이고 몸길이는 7m나 된다. 새끼가 태어나자마자 어미는 새끼를 물 표면으로 밀어 올려 첫 숨을 쉬게 한다. 어미는 새끼를 극진히 보살피며, 1년 동안 가까이 두고 지낸다. 어미는 젖을 먹여 새끼를 키우는데, 특수한 가슴 근육을 움직여 젖을 새끼의 입 안으로 밀어 넣는다. 고래의 젖은 농도가 매우 진하고, 육상 포유류보다 지방, 단백질, 무기염류가 훨씬 더 풍부하다. 이러한 젖을 먹은 새끼는 매우 빨리 자라는데, 흰긴수염고래 새끼는 하루에 몸무게가 약 90kg씩 늘어난다.[178]
대부분의 종에서 번식은 계절적이다. 배란은 수컷의 수정능력과 일치한다. 이러한 주기는 많은 종에서 관찰할 수 있는 계절적 이동과 일반적으로 연관되어 있다. 대부분의 이빨고래는 고정된 짝짓기를 하지 않는다. 많은 종에서 암컷은 한 계절에 여러 파트너를 선택한다. 수염고래는 각 번식 기간 동안 주로 일부일처제를 유지한다.
임신 기간은 9개월에서 16개월까지 다양하다. 쇠돌고래와 흰긴수염고래는 약 11개월 동안 임신한다. 유대류와 단공류를 제외한 모든 포유류와 마찬가지로, 배아는 어미의 혈류에서 영양분을 끌어들이는 기관인 태반에 의해 영양을 공급받는다.
고래류는 보통 새끼 한 마리를 낳는다. 쌍둥이를 낳는 경우, 어미가 두 마리 모두에게 충분한 젖을 생산할 수 없기 때문에 한 마리는 보통 죽는다. 현생 고래류에서 태아는 보통 꼬리부터 나오는 자세를 취한다. 일반적인 믿음과는 달리, 이것은 출산 중 익사 위험을 최소화하기 위한 것이 아니다. 출산의 역학과 태아의 형태와 관련이 있을 가능성이 더 높다.[67] 출산 후 어미는 새끼를 수면으로 데려가 첫 호흡을 하게 한다. 출생 시, 새끼는 성체 길이의 약 1/3이며, 육상 포유류와 비슷하게 독립적으로 활동하는 경향이 있다.
다른 태반 포유류와 마찬가지로 고래류는 발달된 새끼를 낳아 젖샘에서 나오는 젖으로 새끼를 키운다. 젖을 먹일 때 어미는 입술이 없는 새끼의 입에 젖샘 근육을 사용하여 젖을 적극적으로 뿌린다. 이 젖은 일반적으로 지방 함량이 16%에서 46%로 높아 새끼가 빠르게 크고 무게가 증가한다.[58]
많은 소형 고래류의 경우 젖을 먹이는 기간은 약 4개월 지속된다. 대형 종의 경우 1년 이상 지속되며 어미와 새끼 사이에 강한 유대감을 형성한다.
어미는 새끼를 돌보는 일을 전적으로 책임진다. 일부 종에서는 이른바 "이모"들이 가끔 새끼에게 젖을 먹이는 경우도 있다.
이러한 번식 전략은 생존율이 높은 소수의 새끼를 낳게 한다.
4. 4. 수명
고래류는 다른 고등 포유류에 비해 특이할 정도로 오래 사는 것으로 구분된다. 북극고래(''Balaena mysticetus'')와 같은 일부 고래 종은 200년 넘게 살 수 있다. 뼈로 된 내이낭의 나이테를 기준으로, 가장 나이 많은 것으로 알려진 표본은 사망 당시 211세로 밝혀진 수컷이다.[68] 지금까지 알려진 최고령은 대왕고래가 114세, 흰긴수염고래가 110세이며, 소형종이 될수록 최고령은 낮아지는 경향이 있다.[148]4. 5. 사회 행동
많은 고래 종류는 특정한 계절에만 짝짓기를 한다. 짝짓기 형태는 지느러미발로 서로 치거나, 물 위로 뛰어오르거나, 길고 구부러진 지느러미발로 서로 안는 행동을 포함한다. 임신 기간은 보통 10~12개월이며, 한 배에 한 마리의 새끼를 낳지만 드물게 두 마리가 태어나기도 한다. 다른 암컷 한두 마리가 산모의 출산을 돕기도 한다.[178]고래는 태어날 때부터 몸집이 크다. 갓 태어난 흰긴수염고래는 몸무게가 평균 약 1.8ton이고, 몸길이는 7m에 달한다. 어미는 새끼가 태어나자마자 물 표면으로 밀어 올려 첫 숨을 쉬게 한다. 어미는 새끼를 극진히 보살피며, 1년 동안 가까이 두고 젖을 먹여 키운다. 젖은 특수한 가슴 근육의 운동으로 새끼의 입 안으로 밀어 넣어지는데, 농도가 매우 진하고 육상 포유류의 젖보다 지방, 단백질, 무기염류가 훨씬 풍부하다. 이러한 젖을 먹은 새끼는 매우 빨리 자라는데, 흰긴수염고래 새끼는 하루에 몸무게가 약 90kg씩 늘어난다.[178]
대부분의 고래는 무리를 지어 생활하며, 다양한 종류의 소리를 내어 대화한다. 이 소리는 상당히 먼 거리에서도 잘 들을 수 있으며, 무리의 개체를 모으는 데 사용된다. 각각의 울음소리는 연속된 소리로 되어 있으며 20분 정도 지속된 다음 다시 반복된다.[178]
몇몇 고래는 계절에 따라 먹이를 찾아 이동한다. 고래는 사람을 제외하면 천적이 거의 없다. 돌고래 종류인 범고래는 때때로 새끼 고래, 작은 돌고래, 병들거나 약한 긴수염고래를 공격한다. 그러나 대부분의 고래는 사람에게 잡히지 않는 한 늙어 죽을 때까지 산다.[178]
몇몇 고래는 스스로 바닷가에 올라와서 죽는 경우가 있다. 혼자 또는 무리가 함께 해변으로 올라오는 해안 좌초 현상은, 고래가 자기 몸무게에 눌리거나 밀물에 분수공이 막혀 익사하는 결과를 초래한다. 이러한 행동의 이유는 아직 정확히 밝혀지지 않았다.[178]
의식적으로 숨을 쉬는 고래는 수면을 취하지만, 익사 위험 때문에 오랫동안 무의식 상태에 있을 수 없다. 야생 고래의 수면에 대한 지식은 제한적이지만, 사육 상태의 이빨고래는 뇌의 한쪽만 자는 단측구 수면(USWS)을 하는 것으로 알려져 있다. 이는 휴식 중에도 수영, 의식적 호흡, 포식자 및 사회적 접촉 회피를 가능하게 한다.[54]
2008년 연구에 따르면, 향유고래는 일반적으로 낮 동안 수면 중 수직 자세로 얕은 '드리프트 다이빙'(drift-dives)을 하며 잠을 잔다. 이때 고래는 접촉하지 않는 한 지나가는 선박에 반응하지 않아, 잠수 중 잠을 잘 수 있다는 추측이 나왔다.[55]
무리 생활을 하는 고래들은 종종 다른 종과 함께 사냥하기도 한다. 많은 돌고래 종들은 큰 참치 무리와 함께 사냥하며, 큰 물고기 떼를 따라다닌다. 범고래는 무리를 지어 사냥하며 흰돌고래나 심지어 더 큰 고래를 사냥감으로 삼는다. 혹등고래를 비롯한 다른 고래들은 기포 카펫을 공동으로 만들어 크릴이나 플랑크톤을 떼 지어 몰아넣은 후 덮치기도 한다.[58]
고래는 가르치고, 배우고, 협력하고, 계략을 짜고, 슬퍼하는 것으로 알려져 있다.[60]
돌고래와 쇠돌고래와 같은 소형 고래는 복잡한 놀이 행동을 한다. 예를 들어, 물속에 공기를 빠르게 내뿜어 고리 모양을 만들거나, 원을 그리며 헤엄치다가 생성된 나선형 소용돌이에 공기를 주입하여 "거품 고리"를 만든다. 또한 소용돌이 고리를 물어뜯어 여러 개의 거품으로 터뜨리는 것을 즐긴다. 고래는 먹이를 모으는 데 도움이 되도록 거품 그물을 만들기도 한다.[61]

대형 고래도 놀이를 하는 것으로 여겨진다. 남방긴수염고래는 꼬리 지느러미를 물 위로 올리고 상당 시간 동안 같은 자세를 유지하는 "항해" 행동을 보이는데, 이는 놀이의 한 형태로 보이며 아르헨티나와 남아프리카 해안에서 가장 흔하게 관찰된다.[62] 혹등고래도 이러한 행동을 보인다.
자기 인식은 추상적 사고의 징후로 여겨진다. 돌고래는 자기 인식을 가지고 있는 것으로 보인다.[63] 동물의 자기 인식을 위한 가장 일반적인 테스트는 거울 테스트이다. 이 테스트는 동물의 몸에 일시적인 염료를 바르고 거울을 보여준 후, 동물이 자기 인식의 징후를 보이는지 조사하는 방식으로 진행된다.[64]
비판적인 시각에서는 이러한 테스트 결과가 클레버 핸스 효과에 영향을 받기 쉽다고 주장한다. 영장류와 달리 돌고래는 표시나 거울을 직접 만질 수 없기 때문에 자기 인식 행동이 덜 확실하다. 회의론자들은 자기 인식을 나타낸다고 주장하는 행동이 기존의 사회적 행동과 유사하여 연구자들이 이를 사회적 반응으로 오인할 수 있다고 주장한다. 그러나 옹호론자들은 이러한 행동이 다른 개체에 대한 일반적인 반응과 다르다고 반박한다.[64]
1995년, Marten과 Psarakos는 비디오를 사용하여 돌고래의 자기 인식을 테스트했다.[65] 그들은 돌고래에게 자신을 녹화한 실시간 영상, 녹화된 영상, 다른 돌고래의 영상을 보여주었다. 그들은 그들의 증거가 사회적 행동이 아닌 자기 인식을 시사한다고 결론지었다. 이 연구는 복제되지 않았지만, 돌고래는 이후 거울 테스트를 통과했다.[64]
집단적인 의사결정은 많은 시간을 무리 생활로 보내는 여러 고래 종에게 중요한 부분이다.[66] 이러한 의사결정이 어떻게 이루어지는지에 대해서는 아직 알려진 바가 거의 없지만, 연구에 따르면 향유고래 무리에서는 복잡한 합의 의사결정이, 범고래와 같은 다른 종에서는 리더십이 존재한다는 증거가 발견되었다.
4. 6. 이동
몇몇 종류의 고래는 계절에 따라 먹이를 찾아 이동한다. 고래는 사람을 빼고는 천적이 거의 없다. 돌고래 종류인 범고래는 때때로 새끼 고래와 작은 돌고래, 병들거나 약한 긴수염고래를 공격하기도 하지만, 대부분의 고래는 사람에게 잡히지 않는 한 늙어 죽을 때까지 산다.[178]많은 고래 종들은 계절 서식지를 오가기 위해 위도 방향으로 이동하는 회유를 한다. 예를 들어, 귀신고래는 약 16000km를 왕복 이동한다. 이 여정은 바하칼리포르니아의 따뜻한 석호에 있는 겨울 번식지에서 시작하여 알래스카 연안의 베링해, 추크치해, 보퍼트해에 있는 여름 먹이 지역까지 약 8000km~11200km의 해안선을 따라 이동한다.[53]
5. 고래와 인간
사람들은 아주 옛날부터 고래를 사냥하여 고래기름을 연료나 양식으로 사용했다. 20세기에는 포경꾼들이 마구잡이로 고래를 잡아 몇몇 종은 심각한 멸종 위기에 처했다. 그래서 국제포경위원회는 매년 잡을 수 있는 고래 수를 제한하고, 몇 종의 고래는 사냥을 완전히 금지했다.[178]
긴수염고래와 이빨고래는 쓰임새가 달랐다. 긴수염고래의 기름은 주로 식용유와 공업용으로 쓰였고, 가죽, 뼈, 내장에서도 유지를 채취했다. 이빨고래의 경유는 주로 세제, 밀랍, 기계유 등 공업용으로 쓰였으며, 고기는 가공하여 식용으로 이용되었다. 특히, 돌고래라고 불리는 소형 고래는 시장에서 식용으로 판매되기도 했다.
고래는 포경, 어업, 오염, 선박 소음, 기후 변화 등 인간 활동으로 인한 다양한 위협에 직면해 있다. 이 때문에 많은 고래 개체군이 멸종 위기에 처했으며, 일부는 멸종 직전까지 몰리기도 했다. 1931년부터 고래 보호 노력이 시작되었고, 1946년 국제포경위원회(IWC)가 설립되어 고래 개체 수 보호에 힘쓰고 있다. 그러나 일부 국가는 포경 금지 조치를 따르지 않고 상업적 포경을 계속하고 있다.
20세기 후반부터 고래에 대한 인식이 바뀌면서 고래 관광이 인기를 얻고 있다. 고래 관찰은 생태 관광의 하나로 경제적 효과를 올리고 있지만, 고래에게 부담을 줄 수 있다는 우려도 나온다.
5. 1. 문화 속 고래
고래류는 역사를 통틀어 인간 문화에서 중요한 역할을 해왔다. 구석기 시대 암각화, 예를 들어 노르웨이 로도이와 레파의 암각화, 대한민국의 반구대 암각화 등에서 그 모습을 볼 수 있다.[101][102] 오크니 제도의 신석기 시대 유적지인 스카라 브레이에서는 고래 척추뼈로 소스 팬을 만들었다.
고대 그리스의 호메로스는 고래를 케토스(Ketos)라고 불렀는데, 이 용어는 처음에는 모든 큰 해양 동물을 포함했다. 여기서 로마식 고래 명칭인 케투스(Cetus)가 유래되었다. 다른 명칭으로는 암컷 고래를 가리키는 파라이나(phálaina) (아리스토텔레스, 라틴어로는 발라에나(ballaena))와, 수컷 고래를 가리키는 무스쿨루스(musculus, 쥐)가 있었다. 북해 고래는 피세테르(Physeter)라고 불렸는데, 이는 향유고래 ''Physeter macrocephalus''를 의미했다. 아리스토텔레스, 플리니우스, 암브로시우스는 모두 고래가 살아있는 새끼를 낳고 젖을 먹인다고 언급했다. 플리니우스는 분출공과 관련된 문제점을 묘사했고, 암브로시우스는 큰 고래들이 새끼를 보호하기 위해 입 안으로 데려간다고 주장했다.
성경에서 리바이어던은 바다 괴물로서 역할을 한다. 거대한 악어 또는 용과 고래를 특징으로 하는 이 존재는 성경에 따르면 신에 의해 창조되었고[103] 다시 신에 의해 파괴될 것이다.[104][105] 욥기에서 리바이어던은 더 자세하게 묘사되어 있다.[106][107]
요나에는 예언자 요나와 고래에 대한 묘사가 있다. 요나는 니네베 도시에서 도망치던 중 고래에게 삼켜졌다.
돌고래는 고래보다 훨씬 더 자주 언급된다. 아리스토텔레스는 그의 ''동물지''에서 그리스인들의 신성한 동물을 논하고, 수생 동물로서의 역할에 대한 세부 사항을 제시한다. 그리스인들은 돌고래를 "수생 동물의 왕"으로 숭배했고, 물고기라고 불렀다. 돌고래의 지능은 어망에서 벗어나는 능력과 어부들과의 협력에서 분명하게 나타났다.
돌고래는 그리스 신화에 등장한다. 그 지능 때문에 여러 사람을 익사에서 구출했다고 한다. 돌고래는 음악을 좋아한다고 하며, 전설에서 레스보스 메팀나 출신의 음악가 아리온을 구했다고 한다.
돌고래는 포세이돈의 영역에 속하며, 그를 그의 아내 암피트리테에게로 이끌었다. 돌고래는 아폴론, 디오니소스, 아프로디테와 같은 다른 신들과도 관련되어 있다. 그리스인들은 고래와 돌고래 모두에게 자신들의 별자리로 경의를 표했다. 고래자리(케토스, 라틴어 케투스)는 돌고래자리(델피, 라틴어 델피누스)의 남쪽, 황도대의 북쪽에 위치해 있다.
고대 예술에는 크레타의 미노아인을 포함하여 돌고래를 묘사한 작품이 자주 포함되었다. 나중에는 부조, 보석, 램프, 동전, 모자이크 및 묘비에 등장했다. 특히 인기 있는 묘사는 돌고래를 타고 있는 아리온이나 타라스이다. 초기 기독교 미술에서 돌고래는 인기 있는 모티프이며, 때로는 그리스도의 상징으로 사용되었다.
브렌단 성인은 자신의 여행담 ''Navigatio Sancti Brendani''에서 565년에서 573년 사이에 고래와의 만남을 묘사했다. 그는 자신과 동료들이 나무가 없는 섬으로 들어갔는데, 그것은 거대한 고래였고, 그는 이 고래를 야스코니쿠스(Jasconicus)라고 불렀다고 설명했다. 그는 7년 후에 이 고래를 다시 만나 등에 올라 쉬었다.


17세기에 시작된 포경 시대까지, 기록된 대부분의 큰 고래에 대한 묘사는 해변에 좌초된 고래들이었다. 레이먼드 길모어는 1723년부터 1959년까지 엘베 강 하구에서 17마리의 향유고래와 1784년 영국 해안에서 31마리의 향유고래를 기록했다. 1827년에는 대왕고래 한 마리가 오스텐드 해안에 좌초되었다. 고래는 박물관과 순회 전시회에서 관광 명소로 이용되었다.
17세기부터 19세기까지의 포경업자들은 그림으로 고래를 묘사하고 그들의 직업에 대한 이야기를 전했다. 그들은 고래가 해를 끼치지 않는 거대한 동물임을 알고 있었지만, 작살로 찔린 동물들과의 싸움을 묘사했다. 여기에는 거대한 고래, 상어, 바다뱀, 대왕오징어, 문어를 포함한 바다 괴물에 대한 묘사가 포함되었다.
포경 여행에서의 경험을 묘사한 최초의 포경업자들 중에는 영국의 선장 윌리엄 스코어스비가 있으며, 그는 북쪽 수염고래 사냥을 묘사한 ''Northern Whale Fishery''라는 책을 출판했다. 이 책에 이어 1835년 영국의 외과 의사 토마스 비일의 ''Some observations on the natural history of the sperm whale''와 1840년 프레데릭 데벨 베넷의 ''The tale of a whale hunt''이 출판되었다. 고래는 허먼 멜빌의 ''모비 딕''과 쥘 베른의 ''해저 2만리''에서 서술적 문학과 그림으로 묘사되었다.
수염판은 스코틀랜드 국립 박물관에서 양동이의 바닥과 같은 선박 부품을 만드는 데 사용되었다. 바이킹들은 수염판으로 장식된 접시를 만들었는데, 때로는 다리미판으로 해석되기도 했다.
캐나다 북극(동해안)의 푸눅(Punuk)과 툴레 문화(1000~1600년경)에서는[108] 나무 대신 수염판을 사용하여 겨울집의 지붕 지지대로 집을 지었다. 건물의 절반은 땅에 묻혀 있었고 실제 지붕은 아마도 흙과 이끼로 덮인 동물 가죽으로 만들어졌을 것이다.[109]

20세기에는 고래류에 대한 인식이 변화했다. 과학이 고래류가 지능적이고 평화로운 동물임을 밝히면서, 괴물에서 경이로운 생물로 변모했다. 고래와 돌고래 사냥은 고래와 돌고래 관광으로 대체되었다. 이러한 변화는 영화와 소설에 반영되어 있다. 예를 들어, ''플리퍼'' 시리즈의 주인공은 큰돌고래였다. ''씨퀘스트 DSV''(1993–1996) TV 시리즈, ''프리 윌리''와 ''스타트렉 4: 귀환의 항해'' 영화, 그리고 더글러스 애덤스(Douglas Adams)의 ''은하수를 여행하는 히치하이커를 위한 안내서'' 시리즈는 그 예이다.[110]
고래 노래 연구는 인기 있는 앨범 ''혹등고래의 노래''를 탄생시켰다.
인류는 예로부터 고래류 전반을 식용으로 해왔으며, 일본에서도 이미 조몬 시대 전기에 노토 반도의 마와키 유적에서 다수의 돌고래 뼈가 출토된 것으로 보아 이미 돌고래 어업이 시작되었던 것으로 생각된다.[164] 일본에서 포획된 고래류는 고래고기로서 소비되는 것이 기본이며, 거의 모든 부위를 식용으로 한다.[165] 고래를 식용으로 하는 지역은 일본뿐만 아니라 노르웨이와 아이슬란드에도 식용 습관이 있으며, 알래스카와 캐나다의 이누이트 및 카리브해의 여러 나라에서도 고래류의 포획과 식용이 행해지고 있다.[166] 한편 유럽의 많은 국가와 미국 등에서는 고래류를 식용으로 하는 습관이 없고, 주로 고래 기름을 목적으로 포경을 행했다. 고래 기름은 램프용 연료나 양초의 원료로서 수요가 높아 활발하게 생산되었지만, 20세기에 들어서 석유 생산이 활발해짐에 따라 수요가 떨어져 거의 생산되지 않게 되었다.[167] 또한 일본에서는 고기와 기름 이외의 부위도 활발하게 이용되어, 고래 수염은 용수철이나 우산대 등의 공예 재료로, 고래뼈도 공예 재료나 비료로 이용되어 왔다.[168]
5. 2. 포경
포경은 주로 수염고래와 향유고래를 사냥하는 행위이다. 이 활동은 구석기 시대부터 계속되어 왔다.[89]중세 시대에 포경의 이유는 고래고기, 연료로 사용할 수 있는 고래 기름, 그리고 주택 건설에 사용되었던 턱뼈를 포함했다. 중세 말엽, 초기 포경 선단은 보리고래와 같은 수염고래를 목표로 했다. 16세기와 17세기에 네덜란드 선단은 약 300척의 포경선과 18,000명의 선원을 보유하고 있었다.
18세기와 19세기에 수염고래는 특히 고래수염 때문에 사냥되었는데, 이것은 나무를 대체하거나, 코르셋과 크리놀린 치마와 같이 강도와 유연성이 필요한 제품에 사용되었다. 또한, 향유고래에서 발견되는 경랍은 기계 윤활유로, 용연향은 제약 및 향수 산업의 재료로 사용되었다. 19세기 후반에는 폭발성 작살이 발명되어 어획량이 급증했다.
큰 배는 고래잡이꾼들을 위한 "모선"으로 사용되었다. 20세기 전반에는 고래가 원료 공급원으로 매우 중요했다. 이 기간 동안 고래는 집중적으로 사냥되었고, 1930년대에는 3만 마리의 고래가 사냥되었다. 이 수치는 1960년대까지 연간 4만 마리 이상으로 증가했고, 이때 대형 수염고래의 개체 수가 붕괴되었다.
대부분의 사냥 대상 고래는 이제 멸종 위기에 처해 있으며, 일부 대형 고래 개체군은 멸종 직전까지 몰렸다. 대서양과 한국의 회색고래 개체군은 완전히 멸종되었고, 북대서양혹등고래 개체군은 300~600마리로 감소했다. 흰긴수염고래 개체군은 약 14,000마리로 추산된다.
고래 보호를 위한 최초의 노력은 1931년에 시작되었다. 혹등고래(당시 약 100마리)[90]와 같이 특히 멸종 위기에 처한 일부 종은 국제적 보호를 받았고, 최초의 보호 구역이 설립되었다. 1946년에는 고래 개체 수를 감시하고 확보하기 위해 국제포경위원회(IWC)가 설립되었다. 이 기구는 1985년부터 2005년까지 14종의 대형 고래의 상업적 포경을 전 세계적으로 금지했지만, 일부 국가는 이 금지 조치를 준수하지 않고 있다.
혹등고래와 흰긴수염고래와 같은 종의 개체 수는 회복되었지만, 여전히 멸종 위기에 처해 있다. 미국 의회는 해양 포유류 개체 수를 유지하기 위해 1972년 해양 포유류 보호법을 통과시켰다. 이 법은 알래스카에서 매년 수백 마리의 고래를 제외하고는 해양 포유류를 포획하는 것을 금지한다. 일본 포경선은 명목상 과학적 목적으로 다른 종의 고래를 사냥할 수 있다.
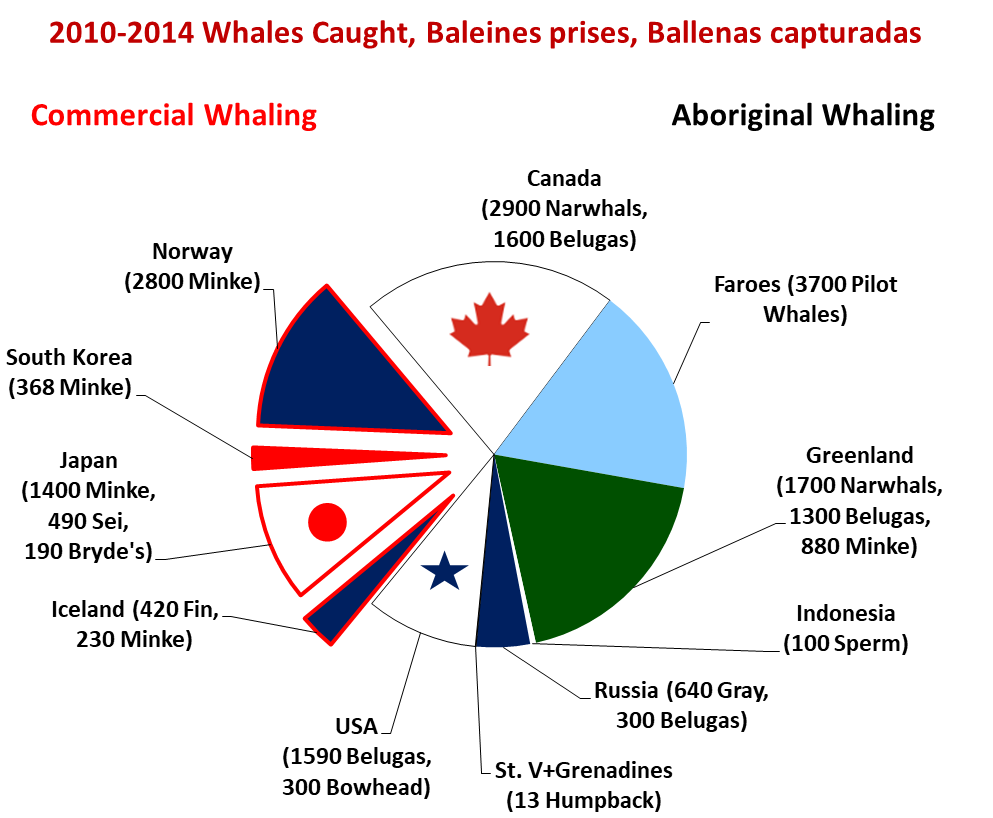
원주민 포경은 여전히 허용된다. 2017년 페로 제도에서는 약 1,200마리의 흑범고래가 포획되었고,[91] 알래스카, 캐나다, 그린란드, 시베리아에서는 매년 약 900마리의 일각돌고래와 800마리의 흰돌고래가 포획된다. 원주민 포경으로 그린란드에서는 매년 약 150마리의 밍크고래, 시베리아에서는 120마리의 회색고래, 알래스카에서는 50마리의 보리고래가 포획되며, 노르웨이가 상업적으로 600마리의 밍크고래, 일본이 300마리의 밍크고래와 100마리의 브라이드고래, 아이슬란드가 최대 100마리의 긴수염고래를 포획하는 것 외에도 말이다.[92] 아이슬란드와 노르웨이는 포경 금지 조치를 인정하지 않고 상업적 포경을 운영한다. 노르웨이와 일본은 포경 금지 조치를 종식시키기 위해 노력하고 있다.
돌고래와 다른 소형 고래류는 돌고래몰이식 사냥으로 알려진 활동에서 때때로 사냥된다. 이것은 보통 배를 사용하여 무리를 몰아 만이나 해변으로 유인하는 방법으로 이루어진다. 다른 배나 그물을 사용하여 바다로 가는 길을 막아 탈출을 막는다. 돌고래는 솔로몬 제도, 페로 제도, 페루 및 일본(가장 잘 알려진 사례)을 포함한 전 세계 여러 곳에서 이러한 방식으로 사냥된다. 돌고래는 주로 고래고기를 위해 사냥되지만, 일부는 돌고래 수족관에 들어간다. 논란에도 불구하고 매년 수천 마리의 돌고래가 돌고래몰이식 사냥으로 포획된다.
인류는 예로부터 고래류 전반을 식용으로 해왔으며, 일본에서도 이미 조몬 시대 전기에 노토 반도의 마와키 유적에서 다수의 돌고래 뼈가 출토된 것으로 보아 이미 돌고래 어업이 시작되었던 것으로 생각된다.[164] 일본에서 포획된 고래류는 고래고기로서 소비되는 것이 기본이며, 거의 모든 부위를 식용으로 한다.[165] 고래를 식용으로 하는 지역은 일본뿐만 아니라 노르웨이와 아이슬란드에도 식용 습관이 있으며, 또한 알래스카와 캐나다의 이누이트 및 카리브해의 여러 나라에서도 고래류의 포획과 식용이 행해지고 있다.[166] 한편 유럽의 많은 국가와 미국 등에서는 고래류를 식용으로 하는 습관이 없고, 주로 고래 기름을 목적으로 포경을 행했다. 고래 기름은 램프용 연료나 양초의 원료로서 수요가 높아 활발하게 생산되었지만, 20세기에 들어서 석유 생산이 활발해짐에 따라 수요가 떨어져 거의 생산되지 않게 되었다.[167] 또한 일본에서는 고기와 기름 이외의 부위도 활발하게 이용되어, 고래 수염은 용수철이나 우산대 등의 공예 재료로, 고래뼈도 공예 재료나 비료로 이용되어 왔다.[168]
고래류는 식량이자 자원으로 유용하기 때문에, 많은 고래류가 서식하는 해역에서는 예로부터 포경이 행해져 왔다. 초기 포경은 해안에 떠밀려 온 개체를 포획하는 정도였을 것으로 생각되지만, 이미 조몬 시대 마와키 유적에서는 돌고래의 몰이식 어업이 행해졌을 것으로 생각된다.[169] 이후 유럽에서는 11세기, 일본에서는 12세기에 포경업이 성립하여, 작살을 이용한 찌르는 방식의 어법으로 포경을 하였으나, 일본에서는 16세기에 그물로 잡는 방식의 포경이 고안되어 주류가 되었다.[170] 20세기에 들어서 포경포로 작살을 쏘는 근대 포경법이 성립하여 전 세계에 퍼졌으나,[171] 효율화된 포경법은 남획을 야기하여 일부 고래류의 개체 수가 급감하였고, 1982년에는 지정된 종의 상업 포경이 금지되었다.[170] 한편, 원주민이 생존 목적으로 하는 원주민 생존 포경은 상업 포경 금지 후에도 조업이 허용되어, 북극해 연안의 미국, 캐나다, 러시아, 그린란드에서는 이누이트 등 원주민에 의해 포경이 계속되고 있으며, 인도네시아 등 여러 지역에서도 포경이 행해지고 있다.[172]
5. 3. 고래 관광
20세기에는 고래류에 대한 인식이 변화했다. 과학이 고래류가 지능적이고 평화로운 동물임을 밝히면서, 괴물에서 경이로운 생물로 변모했다. 고래와 돌고래 사냥은 고래와 돌고래 관광으로 대체되었다. 이러한 변화는 영화와 소설에 반영되어 있다. 예를 들어, ''플리퍼'' 시리즈의 주인공은 큰돌고래였다. ''씨퀘스트 DSV''(1993–1996) TV 시리즈, ''프리 윌리''와 ''스타트렉 4: 귀환의 항해'' 영화, 그리고 더글러스 애덤스(Douglas Adams)의 ''은하수를 여행하는 히치하이커를 위한 안내서'' 시리즈는 그 예이다.[110]고래 노래 연구는 또한 인기 있는 앨범 ''혹등고래의 노래''를 탄생시켰다.
1980년대 후반 이후 고래의 생태를 관찰하는 고래 관찰(ホエールウォッチング)이 전 세계적으로 급속히 인기를 얻게 되었고, 생태 관광(エコツーリズム)의 관점에서도, 또 많은 관광객이 몰려드는 것에 따른 경제적·사회적 관점에서도 기대를 받으며 경제적 효과를 올리고 있다.[175] 한편, 배를 타고 직접 가까이 다가가 관찰하는 행위는 고래류에게 큰 부담을 주는 행위이며, 단기적으로나 장기적으로 영향이 크기 때문에, 법률이나 자율 규제에 의해 어느 정도의 규칙이 만들어진 곳도 있다.[176][177]
소형 고래류는 수족관에서 사육하는 경우가 많으며, 지능이 높은 돌고래나 범고래를 훈련시켜 돌고래쇼를 하는 곳도 많다.[174]


5. 4. 위협과 보존
고래류에 대한 주요 위협은 사람에게서 비롯된다. 직접적인 위협으로는 포경이나 몰이 사냥이 있으며, 간접적인 위협으로는 어업과 오염이 있다.[88]포경은 주로 수염고래와 향유고래를 사냥하는 행위이다. 이 활동은 구석기 시대부터 계속되어 왔다.[89] 중세 시대에는 고래고기, 연료로 사용할 수 있는 고래 기름, 주택 건설에 사용되었던 턱뼈를 얻기 위해 포경을 했다. 중세 말엽, 초기 포경 선단은 보리고래와 같은 수염고래를 목표로 했다. 16세기와 17세기에 네덜란드 선단은 약 300척의 포경선과 18,000명의 선원을 보유하고 있었다.
18세기와 19세기에 수염고래는 특히 수염 때문에 사냥되었는데, 이것은 나무를 대체하거나, 코르셋과 크리놀린 치마와 같이 강도와 유연성이 필요한 제품에 사용되었다. 또한, 향유고래에서 발견되는 경랍은 기계 윤활유로, 용연향은 제약 및 향수 산업의 재료로 사용되었다. 19세기 후반에는 폭발성 작살이 발명되어 어획량이 급증했다.
큰 배는 고래잡이꾼들을 위한 "모선"으로 사용되었다. 20세기 전반에는 고래가 원료 공급원으로 매우 중요했다. 이 기간 동안 고래는 집중적으로 사냥되었고, 1930년대에는 3만 마리의 고래가 사냥되었다. 이 수치는 1960년대까지 연간 4만 마리 이상으로 증가했고, 이때 대형 수염고래의 개체 수가 붕괴되었다.
대부분의 사냥 대상 고래는 이제 멸종 위기에 처해 있으며, 일부 대형 고래 개체군은 멸종 직전까지 몰렸다. 대서양과 한국의 회색고래 개체군은 완전히 멸종되었고, 북대서양혹등고래 개체군은 300~600마리로 감소했다. 흰긴수염고래 개체군은 약 14,000마리로 추산된다.
고래 보호를 위한 최초의 노력은 1931년에 시작되었다. 혹등고래(당시 약 100마리)[90]와 같이 특히 멸종 위기에 처한 일부 종은 국제적 보호를 받았고, 최초의 보호 구역이 설립되었다. 1946년에는 고래 개체 수를 감시하고 확보하기 위해 국제포경위원회(IWC)가 설립되었다. 이 기구는 1985년부터 2005년까지 14종의 대형 고래의 상업적 포경을 전 세계적으로 금지했지만, 일부 국가는 이 금지 조치를 준수하지 않고 있다.
혹등고래와 흰긴수염고래와 같은 종의 개체 수는 회복되었지만, 여전히 멸종 위기에 처해 있다. 미국 의회는 해양 포유류 개체 수를 유지하기 위해 1972년 해양 포유류 보호법을 통과시켰다. 이 법은 알래스카에서 매년 수백 마리의 고래를 제외하고는 해양 포유류를 포획하는 것을 금지한다. 일본 포경선은 명목상 과학적 목적으로 다른 종의 고래를 사냥할 수 있다.
원주민 포경은 여전히 허용된다. 2017년 페로 제도에서는 약 1,200마리의 흑범고래가 포획되었고,[91] 알래스카, 캐나다, 그린란드, 시베리아에서는 매년 약 900마리의 일각돌고래와 800마리의 흰돌고래가 포획된다. 원주민 포경으로 그린란드에서는 매년 약 150마리의 밍크고래, 시베리아에서는 120마리의 회색고래, 알래스카에서는 50마리의 보리고래가 포획되며, 노르웨이가 상업적으로 600마리의 밍크고래, 일본이 300마리의 밍크고래와 100마리의 브라이드고래, 아이슬란드가 최대 100마리의 긴수염고래를 포획하는 것 외에도 말이다.[92] 아이슬란드와 노르웨이는 포경 금지 조치를 인정하지 않고 상업적 포경을 운영한다. 노르웨이와 일본은 포경 금지 조치를 종식시키기 위해 노력하고 있다.
돌고래와 다른 소형 고래류는 돌고래몰이식 사냥으로 알려진 활동에서 때때로 사냥된다. 이것은 보통 배를 사용하여 무리를 몰아 만이나 해변으로 유인하는 방법으로 이루어진다. 다른 배나 그물을 사용하여 바다로 가는 길을 막아 탈출을 막는다. 돌고래는 솔로몬 제도, 페로 제도, 페루 및 일본(가장 잘 알려진 사례)을 포함한 전 세계 여러 곳에서 이러한 방식으로 사냥된다. 돌고래는 주로 고래고기를 위해 사냥되지만, 일부는 돌고래 수족관에 들어간다. 논란에도 불구하고 매년 수천 마리의 돌고래가 돌고래몰이식 사냥으로 포획된다.

돌고래 무리는 종종 큰 참치 떼 근처에 서식한다. 어부들은 이 사실을 알고 돌고래를 찾아 참치를 잡는다. 돌고래는 정기적으로 숨을 쉬기 때문에 참치보다 멀리서도 훨씬 쉽게 발견할 수 있다. 어부들은 돌고래 무리 주위에 수백 미터 너비의 그물을 원형으로 쳐서 참치 떼를 잡을 것을 기대한다. 그물이 끌어당겨지면 돌고래는 물속에서 얽히게 되어 익사한다. 대형 강의 통발 어업은 강돌고래에 위협이 된다.
소형 고래류에게 혼획보다 더 큰 위협은 표적 사냥이다. 동남아시아에서는 해당 지역의 식용 어류가 수출에서 더 높은 수익을 약속하기 때문에 지역 주민들에게 생선 대용으로 판매된다. 지중해에서는 식용 어류에 대한 압력을 줄이기 위해 소형 고래류를 표적으로 한다.[88]
좌초란 고래류가 물 밖으로 나와 해변에 닿는 것을 말한다. 어떤 경우에는 여러 마리의 고래가 함께 좌초하기도 한다. 가장 잘 알려진 것은 파일럿고래와 향유고래의 대규모 좌초이다. 좌초된 고래류는 대개 죽는데, 최대 90MT에 달하는 체중이 폐를 압박하거나 갈비뼈를 부러뜨리기 때문이다. 작은 고래는 체온 유지 능력 때문에 열사병으로 죽을 수도 있다.

원인은 명확하지 않다. 대규모 좌초의 가능한 원인은 다음과 같다.[88]
- 독성 오염물질
- 심각한 기생충 감염 (호흡기, 뇌 또는 중이)
- 감염 (세균성 또는 바이러스성)
- 포식자(인간 포함)로부터의 도피
- 무리 내 사회적 유대감으로 인해 무리가 좌초된 동물을 따라가는 경우
- 지구 자기장의 자연적 이상에 의한 자기 감각의 교란
- 부상
- 선박 교통, 지진 조사 및 군사 소나 실험으로 인한 소음 공해
2000년 이후로 군사용 소나 시험 이후 고래 좌초가 빈번하게 발생했다. 2001년 12월, 미국 해군은 2000년 3월 여러 해양 포유류의 좌초 및 사망에 대해 부분적인 책임을 인정했다. 중간 보고서의 공동 저술자는 일부 해군 함정의 활성 소나에 의해 죽은 동물들이 부상을 입었다고 밝혔다. 일반적으로 여전히 증가하고 있는 수중 소음은 의사소통과 방향 감각을 저해하기 때문에 좌초와 점점 더 관련이 있다.[93]
기후 변화는 주요 풍향 시스템과 해류에 영향을 미치며, 이는 고래류 좌초로 이어진다. 1920년부터 2002년까지 타즈메이니아 해안에서 좌초를 연구한 연구원들은 특정 시간 간격으로 좌초가 더 많이 발생했다는 사실을 발견했다. 좌초가 증가한 해는 심한 폭풍과 관련이 있었는데, 이 폭풍은 해안 가까이에 차가운 해류를 발생시켰다. 영양이 풍부한 차가운 물에서 고래류는 큰 먹이 동물을 기대하기 때문에 차가운 해류를 따라 얕은 물로 이동하며, 이곳에서 좌초 위험이 더 높다. 무리 지어 사는 고래와 돌고래는 아픈 또는 허약한 무리 구성원을 얕은 물로 따라갈 수 있으며, 간조 때 좌초될 수 있다.[94]
중금속, 많은 식물과 곤충 독의 잔류물, 그리고 플라스틱 폐기물 부유물은 생분해되지 않는다. 때때로 고래류는 이러한 유해 물질을 먹이로 오인하여 섭취한다. 결과적으로 동물들은 질병에 더 취약해지고 새끼를 덜 낳습니다.[88]
오존층의 손상은 그로 인한 방사선 때문에 플랑크톤의 번식을 감소시킨다. 이것은 많은 해양 동물의 먹이 공급을 줄이지만, 여과 섭식을 하는 수염고래가 가장 큰 영향을 받는다. 심지어 유영 생물도 집중적인 착취 외에도 방사선에 의해 피해를 입는다.[88]
대기 중 이산화탄소 증가로 인한 해양 산성화로 인해 장기적으로 먹이 공급이 감소하기도 한다. CO2는 물과 반응하여 탄산을 형성하며, 이는 수염고래가 의존하는 동물성 플랑크톤의 먹이 공급원인 탄산칼슘 골격의 형성을 감소시킨다.[88]
군사 및 자원 추출 산업은 강력한 소나와 폭파 작업을 운영한다. 해양 지진 조사는 지구 표면 아래에 무엇이 있는지 보여주는 큰 저주파 소리를 사용한다.[95] 선박 통행 또한 해양의 소음을 증가시킨다. 이러한 소음은 방향 감지와 의사소통을 위한 생체 소나 사용과 같은 고래류의 행동을 방해할 수 있다. 심각한 경우 공황 상태를 유발하여 수면으로 몰아낼 수 있다. 이것은 혈액 가스에 기포를 생성하여 감압병을 유발할 수 있다.[96] 소나를 사용한 해군 훈련은 정기적으로 치명적인 감압으로 인해 좌초되는 고래류를 발생시킨다. 소리는 100km 이상의 거리에서도 방해가 될 수 있다. 피해는 주파수와 종에 따라 다릅니다.
참조
[1]
논문
New protocetid whales from Alabama and Mississippi, and a new Cetacean clade, Pelagiceti
[2]
학회발표
Evolutionary history of the cetaceans: a review
Verlag Dr. Friedrich Pfeil
[3]
논문
A guide to the pronunciation and meaning of cetacean taxonomic names
http://aquaticmammal[...]
[4]
논문
Balancing Requirements for Stability and Maneuverability in Cetaceans
[5]
서적
The Guinness Book of Animal Facts and Feats
[6]
웹사이트
The longest animal alive may be one you never thought of
http://www.bbc.com/e[...]
2016-04-20
[7]
웹사이트
Largest mammal
http://www.guinnessw[...]
[8]
웹사이트
World Cetacea Database
http://www.marinespe[...]
2020-00-00
[9]
논문
Investigating the Roles of Cetaceans in Marine Ecosystems - An overview
https://www.research[...]
2004-00-00
[10]
서적
Current State of Knowledge of Cetacean Threats, Diversity, and Habitats in the Pacific Islands Region
http://uk.whales.org[...]
Whale and Dolphin Conservation Society
[11]
논문
Responsible Practices for Minimizing and Monitoring Environmental Impacts of Marine Seismic Surveys with an Emphasis on Marine Mammal
2013-00-00
[12]
뉴스
China's Rare River Dolphin Now Extinct, Experts Announce
http://news.national[...]
National Geographic Society
2006-12-14
[13]
논문
Influence of Alignment on the mtDNA Phylogeny of Cetacea: Questionable Support for a Mysticeti/Physeteroidea Clade
1998-06-01
[14]
뉴스
Ungulate taxonomy
[15]
서적
The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea
https://books.google[...]
Springer
2013-11-11
[16]
서적
Reproductive Biology and Phylogeny of Cetacea: Whales, Porpoises and Dolphins
https://books.google[...]
CRC Press
2007-00-00
[17]
서적
Whales of the World
https://books.google[...]
Brill Archive
1988-01-01
[18]
논문
A trade-off between precopulatory and postcopulatory trait investment in male cetaceans
2015-00-00
[19]
논문
Phylogeny of the Beaked Whale Genus ''Mesoplodon'' (Ziphiidae: Cetacea) Revealed by Nuclear Introns: Implications for the Evolution of Male Tusks
2008-00-00
[20]
웹사이트
How ancient whales lost their legs, got sleek and conquered the oceans
http://www.eurekaler[...]
University of Florida
2006-05-22
[21]
논문
Anatomy of nasal complex in the southern right whale, Eubalaena australis (Cetacea, Mysticeti)
2015-00-00
[22]
서적
Development of the Cetacean Nasal Skull
https://books.google[...]
Springer
1999-01-29
[23]
웹사이트
Sperm Whales brain size
http://www.nmfs.noaa[...]
NOAA Fisheries – Office of Protected Resources
[24]
잡지
Are whales smarter than we are?
http://blogs.scienti[...]
[25]
논문
The prefrontal cortex of the bottlenose dolphin (Tursiops truncatus Montagu, 1821): a tractography study and comparison with the human
2023-00-00
[26]
웹사이트
Allometry
http://pages.ucsd.ed[...]
University of California San Diego
[27]
서적
Evolutionary Biology: Convergent Evolution, Evolution of Complex Traits
Springer
2016-00-00
[28]
논문
Dendritic architecture of the Von Economo neurons
[29]
논문
Intuition and autism: a possible role for Von Economo neurons
[30]
논문
Structure of the cerebral cortex of the humpback whale, ''Megaptera novaeangliae'' (Cetacea, Mysticeti, Balaenopteridae)
[31]
웹사이트
Why do whale and dolphin tails go up and down?
https://us.whales.or[...]
[32]
웹사이트
Whales, dolphins and porpoises
https://www.dcceew.g[...]
Australian Government Department of Climate Change, Energy, the Environment and Water
[33]
서적
Comparative Physiology of the Vertebrate Digestive System
https://books.google[...]
University of Cambridge
[34]
논문
Seawater Consumption and Water Flux in the Common Dolphin ''Delphinus delphis''
[35]
서적
The Works of John Hunter, F.R.S.
https://books.google[...]
Cambridge University
2015-03-26
[36]
서적
Encyclopedia of Marine Mammals
https://books.google[...]
Elsevier Science
2017-11-27
[37]
서적
Reproductive Biology and Phylogeny of Cetacea: Whales, Porpoises and Dolphins
2007
[38]
잡지
Reproductive Thermoregulation in Marine Mammals
https://uncw.edu/mms[...]
1998
[39]
서적
The Emergence of Whales
Springer US
1998
[40]
학술지
On the Olfactory Anatomy in an Archaic Whale (Protocetidae, Cetacea) and the Minke Whale Balaenoptera acutorostrata (Balaenopteridae, Cetacea)
2013
[41]
웹사이트
Cetacea
https://school.eb.co[...]
Encyclopædia Britannica, Inc.
[42]
웹사이트
Guiana Dolphins Can Use Electric Signals to Locate Prey
http://news.sciencem[...]
American Association for the Advancement of Science (AAAS)
2011-07
[43]
서적
Encyclopedia of Marine Mammals
Academic Press
[44]
서적
The Evolutionary Biology of Hearing
Springer
[45]
서적
Encyclopedia of Marine Mammals
https://books.google[...]
Academic Press
[46]
학술지
Estimating the number of sperm whale (Physeter macrocephalus) individuals based on grouping of corresponding clicks
https://www.research[...]
[47]
학술지
Wonky whales: the evolution of cranial asymmetry in cetaceans
2020-07-10
[48]
학술지
Wonky whales: the evolution of cranial asymmetry in cetaceans
2020-07-10
[49]
학술지
Structure and function in whale ears
http://csi.whoi.edu/[...]
[50]
학술지
Comparative chromosome studies in Cetacea
[51]
학술지
Independent adaptation to riverine habitats allowed survival of ancient cetacean lineages
2000
[52]
학술지
Genetic structure of cetacean populations in sympatry, parapatry, and mixed assemblages: implications for conservation policy
[53]
웹사이트
Gray Whale Migration
https://journeynorth[...]
[54]
학술지
Sleep behaviour: sleep in continuously active dolphins
2006-06
[55]
학술지
Stereotypical resting behavior of the sperm whale
2008-01
[56]
학술지
Experimental investigations on the respiratory function in diving mammals and birds
[57]
학술지
Structure and biomechanical properties of the trachea of the striped dolphin Stenella coeruleoalba: Evidence for evolutionary adaptations to diving
[58]
서적
Cetacean Societies: Field Study of Dolphins and Whales
University of Chicago
[59]
학술지
Cetacean vocal learning and communication
2014
[60]
뉴스
Watching Whales Watching Us
https://www.nytimes.[...]
2009-07-08
[61]
학술지
Underwater components of humpback whale bubble-net feeding behaviour
[62]
서적
Whales, Dolphins and Porpoises
[63]
웹사이트
Elephant Self-Awareness Mirrors Humans
http://www.livescien[...]
Live Science
2006-10-30
[64]
뉴스
Mirror test
https://www.nytimes.[...]
2001-05
[65]
학술지
Using Self-View Television to Distinguish between Self-Examination and Social Behavior in the Bottlenose Dolphin (''Tursiops truncatus'')
1995-06
[66]
학술지
Collective Decision-making in Aquatic Mammals
2023-10
[67]
학술지
Heads or tails first? Evolution of fetal orientation in ichthyosaurs, with a scrutiny of the prevailing hypothesis
2023-04-18
[68]
학술지
Age and growth estimates of bowhead whales (''Balaena mysticetus'') via aspartic acid racemization
[69]
서적
Ecology of Whale Falls at the Deep-Sea Floor
http://www.soest.haw[...]
2003
[70]
학술지
Three-year investigations into sperm whale-fall ecosystems in Japan
2007-02-16
[71]
논문
Brucella ceti and Brucellosis in Cetaceans
[72]
논문
New Protocetid Whale from the Middle Eocene of Pakistan: Birth on Land, Precocial Development, and sexual dimorphism
[73]
논문
Farewell to life on land - thoracic strength as a new indicator to determine paleoecology in secondary aquatic mammals
2016-07-10
[74]
논문
Phylogenetic aspects of Cetacean origins: A morphological perspective
[75]
웹사이트
First intact fossil of prehistoric whale discovered in Wadi Al-Hitan
https://www.iucn.org[...]
International Union for Conservation of Nature
2015-06-09
[76]
논문
Whales originated from aquatic artiodactyls in the Eocene epoch of India
http://repository.ia[...]
[77]
논문
More DNA support for a Cetacea/Hippopotamidae clade: the blood-clotting protein gene gamma-fibrinogen
1997-05-01
[78]
논문
The position of Hippopotamidae within Cetartiodactyla
[79]
웹사이트
Scientists find missing link between the dolphin, whale and its closest relative, the hippo
http://www.innovatio[...]
2005-01-25
[80]
논문
A higher-level MRP supertree of placental mammals
[81]
논문
The placental mammal ancestor and the post-K-Pg radiation of placentals
[82]
논문
Resolving conflict in eutherian mammal phylogeny using phylogenomics and the multispecies coalescent model
[83]
논문
Phylogenomic datasets provide both precision and accuracy in estimating the timescale of placental mammal phylogeny
[84]
논문
Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation
[85]
논문
A phylogenetic blueprint for a modern whale
https://www.montclai[...]
2015-09-04
[86]
웹사이트
Small cetaceans
https://iwc.int/smal[...]
International Whaling Commission
[87]
웹사이트
Lives of Whales
https://iwc.int/live[...]
International Whaling Commission
[88]
서적
Current State of Knowledge of Cetacean Threats, Diversity, and Habitats in the Pacific Islands Region
http://uk.whales.org[...]
Whale and Dolphin Conservation Society
[89]
웹사이트
Whaling - Commercial, Industrial, Japan Britannica
https://www.britanni[...]
2024-10-11
[90]
웹사이트
A History of the International Whaling Commission (IWC)
https://wwf.panda.or[...]
[91]
웹사이트
Hagar & seyðamark
http://heimabeiti.fo[...]
[92]
웹사이트
Total Catches
https://iwc.int/tota[...]
[93]
논문
Whale deaths caused by US Navy's sonar
[94]
논문
Periodic variability in cetacean strandings: links to large-scale climate events
2005-06-22
[95]
논문
Responsible Practices for Minimizing and Monitoring Environmental Impacts of Marine Seismic Surveys with an Emphasis on Marine Mammal
2013
[96]
서적
Europe Oceans 2005
Oceans 2005–Europe
[97]
웹사이트
The History of Animals, Book VIII
https://penelope.uch[...]
[98]
서적
Historiae animalium
http://etext.lib.vir[...]
2008-09-06
[99]
논문
An account of the dissection of a porpess
[100]
논문
Studies for the construction of epicranialen airway when porpoise (Phocoena phocoena Linnaeus, 1758)
[101]
웹사이트
PCAS Quarterly - Rock Art on the Channel Islands of California
http://www.pcas.org/[...]
[102]
뉴스
BBC News - Rock Art Hints at Whaling Origins
http://news.bbc.co.u[...]
2004-04-20
[103]
성경
[104]
성경
[105]
성경
[106]
성경
[107]
성경
[108]
논문
The circumpolar zone
[109]
논문
The Role of Architectural utility in the formation of archaeological Whale Bone Assemblages
[110]
웹사이트
Movie Retriever: Whales
http://www.movieretr[...]
[111]
웹사이트
The Whales, New York Tribune, August 9, 1861
http://chnm.gmu.edu/[...]
1861-08-09
[112]
웹사이트
Beluga Whales in Captivity: Hunted, Poisoned, Unprotected
http://webpages.char[...]
Canadian Marine Environment Protection Society
2006
[113]
웹사이트
Beluga (Delphinapterus leucas) Facts – Distribution – In the Zoo
http://www.waza.org/[...]
World Association of Zoos and Aquariums
[114]
서적
Whales
https://archive.org/[...]
Facts on File
[115]
웹사이트
Conservation Plan for Southern Resident Killer Whales (Orcinus orca)
http://www.nwr.noaa.[...]
National Marine Fisheries Service (NMFS) Northwest Regional Office
[116]
웹사이트
Killer Controversy: Why Orcas Should No Longer Be Kept in Captivity
http://www.hsi.org/a[...]
Humane Society International and the Humane Society of the United States
[117]
뉴스
Whale Attack Renews Captive Animal Debate
https://www.cbsnews.[...]
2010-03-01
[118]
서적
Animal Ethics Reader
Psychology Press
[119]
웹사이트
Orcas in Captivity – A look at killer whales in aquariums and parks
http://www.orcahome.[...]
2009-11-23
[120]
웹사이트
Chapter I: Space requirements
http://www.ecfr.gov/[...]
[121]
웹사이트
In the Wake of Blackfish – Is it Time to Retire the Last Killer Whale Whose Capture Was Shown in the Film?
https://www.huffpost[...]
2013-10-29
[122]
웹사이트
Shamu Goes Out With the Tide: SeaWorld CEO On Its Abrupt Change – And What Comes Next
https://www.forbes.c[...]
2016-03-24
[123]
서적
Dolphins, Porpoises, and Whales of the World: the IUCN Red Data Book
https://portals.iucn[...]
[124]
논문
Growth of two gray whale calves
http://www.aquaticma[...]
[125]
웹사이트
Appendices I, II and III
https://cites.org/en[...]
2021-06-22
[126]
서적
Order Cetacea
http://www.departmen[...]
Johns Hopkins University Press
[127]
서적
クジラとは?
平凡社
[128]
논문
世界哺乳類標準和名目録
日本哺乳類学会
[129]
논문
哺乳類の日本語分類群名,特に目名の取扱いについて —文部省の“目安”にどう対応するか—
日本哺乳類学会
[130]
논문
神奈川県におけるサラワクイルカ座礁の初記録
神奈川県立生命の星・地球博物館
[131]
웹사이트
Indo-European Etymology Database
http://starling.rine[...]
[132]
서적
説文解字
[133]
서적
中國漢字文物大系
大象出版社
[134]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[135]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[136]
서적
イルカ概論 日本近海産小型鯨類の生態と保全
東京大学出版会
2019-02-15
[137]
서적
シャチ学
東海教育研究所
2021-07-30
[138]
기타
[139]
기타
[140]
서적
シャチ学
東海教育研究所
2021-07-30
[141]
서적
鯨類学
東海大学出版会
2008-05-20
[142]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[143]
서적
鯨類学
東海大学出版会
2008-05-20
[144]
서적
続 イルカ・クジラ学
東海大学出版部
2015-04-20
[145]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[146]
서적
鯨類学
東海大学出版会
2008-05-20
[147]
서적
イルカ概論 日本近海産小型鯨類の生態と保全
東京大学出版会
2019-02-15
[148]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[149]
서적
鯨類学
東海大学出版会
2008-05-20
[150]
서적
イルカ概論 日本近海産小型鯨類の生態と保全
東京大学出版会
2019-02-15
[151]
논문
哺乳類の高次分類群および分類階級の日本語名称の提案について
2003
[152]
서적
Suborder Whippomorpha — Hippopotamuses, Cetaceans
Bloomsbury Publishing
2013
[153]
서적
Introduction
Lynx Edicions
2020
[154]
웹사이트
捕鯨を取り巻く状況
https://www.jfa.maff[...]
2024-03-05
[155]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[156]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[157]
웹사이트
捕鯨
https://www.mofa.go.[...]
2024-03-05
[158]
간행물
絶滅のおそれのある野生動植物の種の保存に関する法律施行令 別表第二 国際希少野生動植物種
http://www.env.go.jp[...]
[159]
서적
日本の希少な野生水生生物に関するデータブック
財団法人自然環境研究センター
2000
[160]
기타
[161]
방송
そうだったのか!池上彰の学べるニュース
2010-07-21
[162]
기타
[163]
웹사이트
ねこの目通信 研究室訪問 粕谷研究室
http://www.animalweb[...]
[164]
웹사이트
イルカ漁のムラ
http://www.mawakiise[...]
2024-03-03
[165]
서적
FOOD'S FOOD 新版 食材図典 生鮮食材編
小学館
2003-03-20
[166]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[167]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[168]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[169]
서적
イルカ概論 日本近海産小型鯨類の生態と保全
東京大学出版会
2019-02-15
[170]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[171]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[172]
서적
クジラ・イルカの疑問50
成山堂書店
2018-06-28
[173]
웹사이트
鯨とともに生きる
https://japan-herita[...]
2024-03-05
[174]
서적
シャチ学
東海教育研究所
2021-07-30
[175]
서적
鯨類学
東海大学出版会
2008-05-20
[176]
서적
鯨類学
東海大学出版会
2008-05-20
[177]
웹사이트
ホエールウォッチングについて今知っておきたいこと
https://natgeo.nikke[...]
2024-03-05
[178]
글로벌2 # 글로벌2 타입에 대한 정보가 부족하여 그대로 유지합니다. 추가 정보가 필요합니다.
고래
[179]
논문
Relationships of Cetacea (Artiodactyla) Among Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution
2009
[180]
논문
A phylogenetic blueprint for a modern whale
https://www.montclai[...]
2013-02
[181]
뉴스
고래 `대변`이 알고보면 바다의 보물?
https://web.archive.[...]
매일경제
2010-10-15
[182]
뉴스
오늘의 호기심 고래가 물을 뿜는 이유
https://www.joongang[...]
중앙일보
[183]
웹사이트
고래가 물을 뿜는 이유
http://lg-sl.net/pro[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com