인류 Y-염색체 DNA 하플로그룹
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
- 1. 개요
- 2. 명명 규칙
- 3. 계통 발생 구조
- 4. 주요 Y-DNA 하플로그룹
- 4.1. 하플로그룹 A, B
- 4.2. 하플로그룹 C (M130)
- 4.3. 하플로그룹 D (CTS3946)
- 4.4. 하플로그룹 E (M96)
- 4.5. 하플로그룹 F (M89)
- 4.6. 하플로그룹 G (M201)
- 4.7. 하플로그룹 H (M69)
- 4.8. 하플로그룹 I (M170)
- 4.9. 하플로그룹 J (M304)
- 4.10. 하플로그룹 K (M9)
- 4.11. 하플로그룹 L, T (K1)
- 4.12. 하플로그룹 K2 (K-M526)
- 4.13. 하플로그룹 K2a, K2a1, NO & NO1
- 4.14. 하플로그룹 N (M231)
- 4.15. 하플로그룹 O (M175)
- 4.16. 하플로그룹 K2b1, M & S
- 4.17. 하플로그룹 P (K2b2)
- 4.18. 하플로그룹 Q (M242)
- 4.19. 하플로그룹 R (M207)
- 5. 한국과 관련된 주요 하플로그룹
- 6. 하플로그룹 연대 추정
- 참조
1. 개요
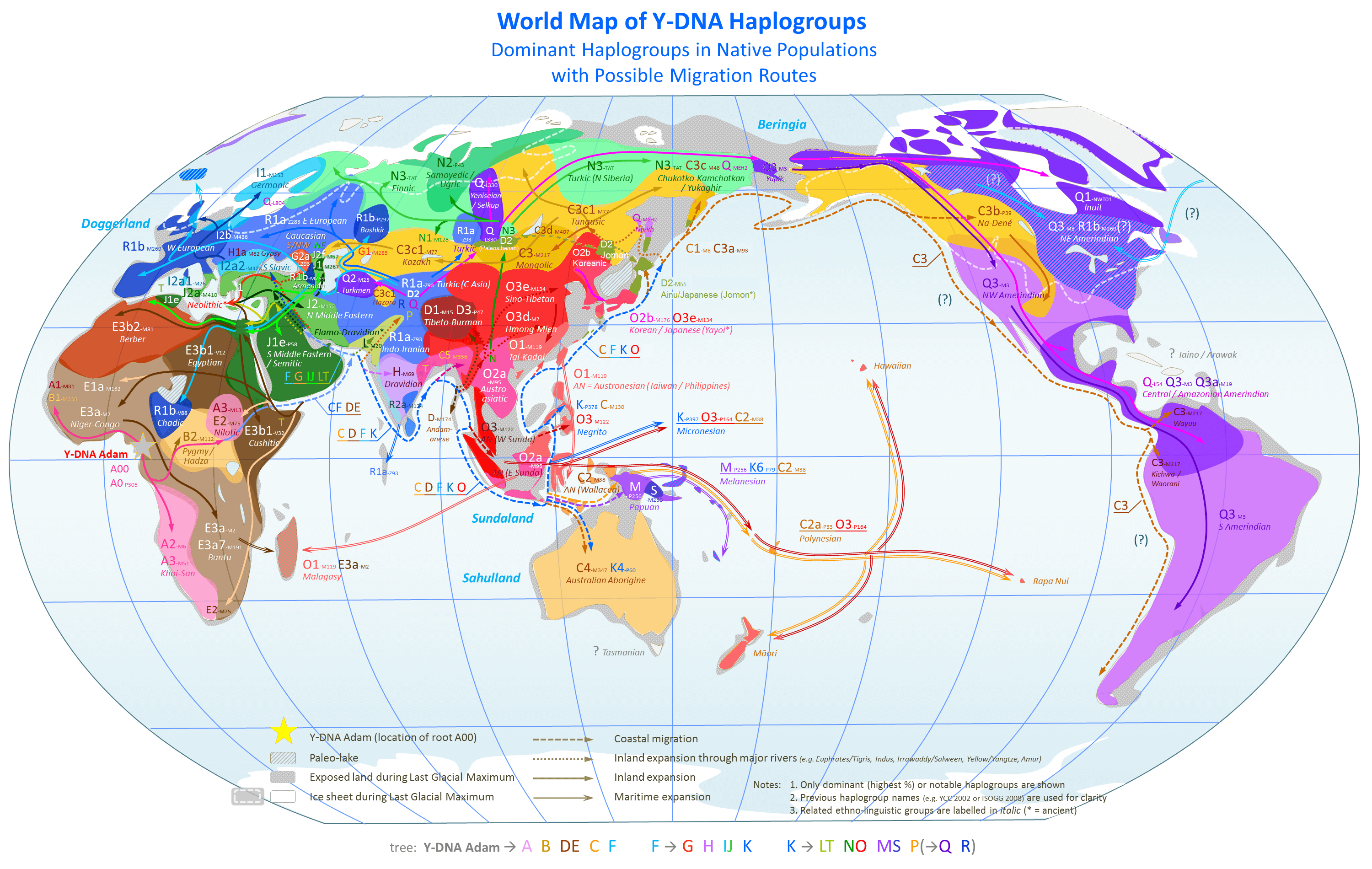
인류 Y-염색체 DNA 하플로그룹은 Y 염색체의 DNA 유전자 표지자에 의해 정의되며, 부계 유전을 통해 전해지는 유전적 계통을 나타낸다. Y-DNA 하플로그룹은 Y 염색체 컨소시엄(YCC)의 명명법에 따라 대문자(A~T)로 주요 하플로그룹을 표시하고, 숫자와 소문자를 조합하여 하위 그룹을 나타낸다. 하플로그룹은 계통 발생 트리를 형성하며, 주요 하플로그룹으로는 아프리카 기원의 A, B, BT, CT, DE, E, CF, C, F, IJK, IJ, I, J, K 등이 있으며, 각 하플로그룹은 특정 지역과 민족 집단에 주로 분포한다. 한국인에게는 O, D, C, N 계통 하플로그룹이 주류를 이루며, 하플로그룹의 연대 추정 결과 Y 염색체 최신 공통 조상은 약 23만 6천 년 전 아프리카에서 기원한 것으로 알려져 있다.

더 읽어볼만한 페이지
- 인류 Y-DNA 하플로그룹 - 하플로그룹 K (Y-DNA)
하플로그룹 K는 약 47,000~50,000년 전 중동 또는 중앙아시아에서 기원한 것으로 추정되는 Y-염색체 DNA 하플로그룹으로, 하위 계통인 L, T, NO, M, S, P 등을 통해 유라시아, 오세아니아, 아메리카 대륙 등 전 세계에 분포한다. - 인류 Y-DNA 하플로그룹 - Y염색체 아담
Y염색체 아담은 현존하는 모든 인류의 부계 혈통을 따라 올라갈 때 만나는 가장 최근의 공통 조상을 지칭하는 비유적인 용어이며, 단 한 명의 남성이 아닌 Y염색체에 돌연변이가 발생하기 이전의 남성 집단을 의미한다.
2. 명명 규칙
Y 염색체 컨소시엄(YCC)은 Y-DNA 하플로그룹을 명명하는 시스템을 개발했다. 이 시스템은 대문자 A부터 T까지를 사용하여 주요 하플로그룹을 나타내고, 숫자와 소문자를 조합하여 하위 그룹을 나타낸다. 예를 들어, 'A'는 가장 오래된 주요 하플로그룹 중 하나를 나타내며, 'A1', 'A1a', 'A1b' 등은 A 하플로그룹 내의 하위 그룹을 나타낸다.[10] 이러한 명명법을 YCC longhand 명명법이라고 한다.
Y-DNA 하플로그룹은 Y염색체 아담을 기원으로 하는 계통수를 형성하며, 주요 분기는 다음과 같다.[82]
Y 염색체의 최신 공통 조상(Y-MRCA, 비공식적으로는 Y 염색체 아담)은 현재 살아있는 모든 인간이 부계로 자손을 남긴 최신 공통 조상이다. Y 염색체를 가진 아담은 약 23만 6000년 전에 아프리카에 살았던 것으로 추정된다.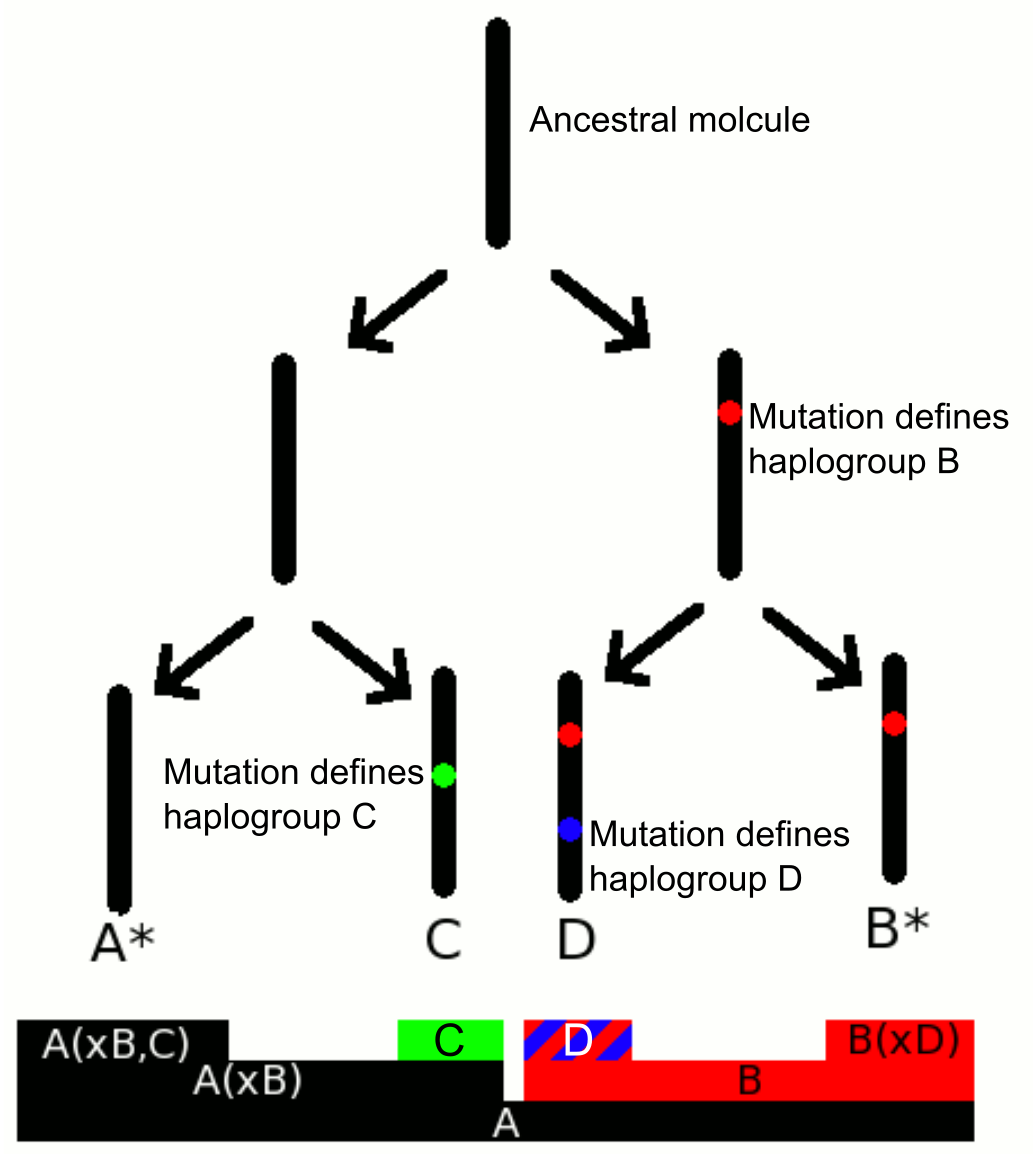
YCC 축약 명명법은 주요 하플로그룹의 첫 글자와 해당 하플로그룹을 정의하는 종말 SNP (Y 염색체 계통 발생 트리의 가장 아래에 있는 SNP)의 이름을 대시(-)로 연결하여 나타낸다.[10] 예를 들어, 하플로그룹 E는 M40이라는 SNP에 의해 정의되므로, 축약 명명법으로는 E-M40으로 표현할 수 있다.
SNP의 발견 및 테스트가 증가함에 따라 Y 염색체 계통 발생 트리가 확장되면서, 하플로그룹 명명법도 계속 변화하고 있다.[2] 이로 인해 자료마다 명명법이 일관되지 않은 경우가 발생하기도 한다. 또한, longhand 명명법이 점점 복잡해짐에 따라 더 간단한 축약 명명법의 사용이 증가하는 추세이다.[11]
3. 계통 발생 구조
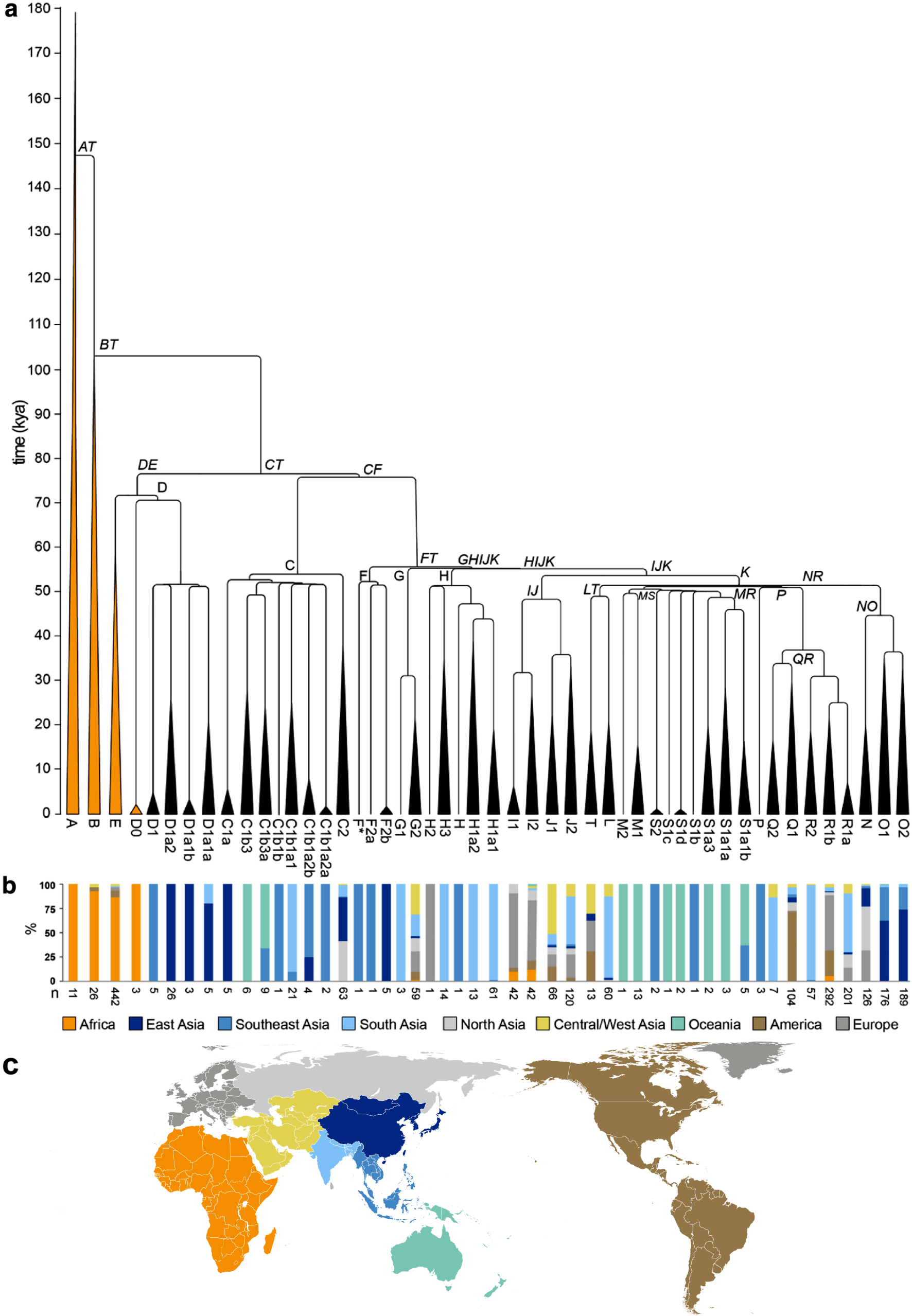
;Y-DNA 하플로그룹의 계통 발생 트리[12]
Y 염색체의 최신 공통 조상(Y-MRCA, 비공식적으로는 Y염색체 아담으로 알려져 있음)은 현재 살아있는 모든 인간이 부계로 자손을 남긴 최신 공통 조상(MRCA)이다. Y 염색체를 가진 아담은 약 23만 6000년 전에 아프리카에 살았던 것으로 추정된다.
대부분의 유라시아인 남성(아프리카 이외의 인구 출신 남성)은 6만 9000년 전에 아프리카에 살았던 남성(Haplogroup_CT)의 자손이다. 다른 주요 병목 현상은 약 5만 년 전과 5천 년 전에 발생했으며, 그 후 대부분의 유라시아인 조상은 5만 년 전에 살았던 4명의 조상(E-M168의 자손)으로 거슬러 올라갈 수 있었다.
언어학상의 어족의 분포와 상관관계가 높지만(부계 언어 가설[70]), 외견상의 인종 구분과는 다른 패턴이 적지 않게 있다(이는 유전자의 계통과 집단의 계통이 다른 불완전 유전자 계통 분리에 의한 것이다).
4. 주요 Y-DNA 하플로그룹
대부분의 유라시아인 남성(아프리카 이외의 인구 출신 남성)은 6만 9000년 전에 아프리카에 살았던 남성(CT)의 자손이다. 약 5만 년 전과 5천 년 전에 발생한 주요 병목 현상 이후, 대부분의 유라시아인 조상은 5만 년 전에 살았던 4명의 조상(E-M168)으로 거슬러 올라갈 수 있었다.
언어학상의 어족의 분포와 상관관계가 높지만(부계 언어 가설[70]), 외견상의 인종 구분과는 다른 패턴이 적지 않게 있다(이는 유전자의 계통과 집단의 계통이 다른 불완전 유전자 계통 분리에 의한 것이다).
주요 Y-DNA 하플로그룹은 다음과 같다.
사키야 미쓰루는 인류의 Y 염색체 하플로그룹이 아프리카 대륙을 벗어난 후 이란 부근을 기점으로 하여 '''남쪽 루트''', '''북쪽 루트''', '''서쪽 루트'''의 3개 루트로 확산되었다고 주장한다[73][74]。
2020년에 발표된 연구에 따르면, 7만 년 ~ 5만 5천 년 전에 일어난 D 계통, C 계통, F 계통의 유전자를 가진 집단의 아프리카 대륙 출신에 이어, 새로운 이민을 제외한 모든 비 아프리카인 남성의 Y 염색체 계통은 5만 5천 년 ~ 5만 년 전에 동아시아/동남아시아에서 확산되었다. 이 기층이 된 계통은 빠르게 유라시아 전체에 걸쳐 확산되었고, 후에 동남아시아에서 다양한 계통으로 분기되었으며, 그 후 약 5만 5천 년 ~ 5만 년 전에 서쪽으로 확산되어, 선주민의 Y 염색체 계통과 대체되었다고 한다[75]。
4. 1. 하플로그룹 A, B
하플로그룹 A는 아프리카에 주로 분포하며, 코이산인, 에티오피아인, 나일 제족과 관련이 있다.[73] 하플로그룹 B는 아프리카에 분포하며, 피그미인과 하드자베인과 관련이 있다.[73]
하플로그룹 A는 모든 현대 부계 하플로그룹이 파생된 NRY(Y 염색체의 비재조합 영역)의 매크로하플로그룹으로, 아프리카에 드물게 분포하며 남서부의 코이산족과 나일강 계곡 북동부의 나일족에게 집중되어 있다. BT는 A 하플로그룹의 하위 분류이며, 더 정확하게는 A1b 계통(Cruciani 외, 2011의 A2-T)의 하위 분류이다.
4. 2. 하플로그룹 C (M130)
하플로그룹 C (M130, xM217)는 오세아니아, 오스트레일리아 원주민, 마오리족, 폴리네시아인에게서 발견된다.[73][74]하플로그룹 C는 크게 하플로그룹 C1 (F3393/Z1426)과 하플로그룹 C2 (M217)로 나뉜다.
하플로그룹 C1은 다시 다음과 같이 세분화된다.
- C1a1 (M8): 일본 고유 하플로그룹으로, 현대 일본인 남성의 약 5~6%에게서 발견된다. 조몬인에 기원한 것으로 추정되지만, 조몬 시대 인골에서 검출된 사례는 아직 발표되지 않았다. 아오야기카미데라지 유적의 야요이 시대 인골이나 다카마쓰차우스야마 고분에 매장된 인골에서 검출된 예는 있다.
- C1a2 (V20): 유럽, 북아프리카, 아르메니아, 네팔, 카빌인(베르베르인 또는 북부 셈족) 등에서 드물게 발견된다. 크로마뇽인과 관련이 있다.
- C1b2 (B477): 오세아니아에서 발견된다.
- C1b2a는 뉴기니 섬 선주민(파푸아인 또는 파푸아 아족)
- C1b2b는 오스트레일리아 선주민(애보리지니 또는 오스트레일리아인종)과 관련이 있다.
하플로그룹 C2 (M217)는 동아시아, 인도차이나 반도, 중앙아시아에서 발견된다.[73][74] 카자흐 스텝, 몽골 고원, 시베리아, 극동 러시아, 북아메리카에서 고빈도로 발견되며, 카자흐인(투란인종 또는 투란인종), 부랴트인, 에벤키 등의 알타이어족, 코랴크인, 니브흐 등의 고시베리아어족(고층 시베리아인종), 아파치족 등의 나데네어족(북태평양 인디언 또는 알류샨인종)과 관련이 있다. 중국 화북, 한반도에서는 중빈도로 발견된다. 중국 화남, 일본, 베트남, 타이족 등에서도 저빈도로 발견된다.
4. 3. 하플로그룹 D (CTS3946)
하플로그룹 D (M174)는 티베트, 일본, 안다만 제도에서 발견된다.하플로그룹 D1a2a (M64.1)는 주로 일본에서 발견되며, 아이누와 오키나와 주민에게서 높은 빈도로 나타난다. 조몬인과의 관련성이 유력하게 제시된다.
4. 4. 하플로그룹 E (M96)
하플로그룹 E (M96)는 주로 아프리카에서 발견된다.하플로그룹 E는 크게 하플로그룹 E1b1a (V38)와 하플로그룹 E1b1b (M215)로 나뉜다. 하플로그룹 E1b1a (V38)는 서아프리카와 그 주변 지역에서 주로 발견되며, 이전에는 E3a로 불렸다. 하플로그룹 E1b1b (M215)는 동아프리카, 북아프리카, 중동, 지중해, 발칸 등에서 발견되며, 이전에는 E3b로 불렸다.[73]
사키야 미쓰루는 인류의 Y 염색체 하플로그룹이 아프리카 대륙을 벗어난 후 이란 부근을 기점으로 확산되었을 때, 하플로그룹 E는 비 아프리카 대륙 출신 그룹에 속한다고 주장한다.[74]
하플로그룹 E1b1a는 사하라 사막 이남의 아프리카, 멜라노 아프리카인종(콩고이드), 반투계 민족 등의 니제르콩고어족과 관련이 있다. 한편, E1b1b는 북아프리카, 아라비아 반도, 이집트 민족(동부 셈족) 등 아프리카아시아어족과 관련이 있다.
4. 5. 하플로그룹 F (M89)
하플로그룹 F는 사하라 사막 이남 아프리카를 제외한 전 세계 인구의 약 90%에서 발견된다.[17]F*(F xF1, F2, F3)는 스리랑카와 남인도 남성의 10%, 파키스탄 남성의 5%에서 보고되었으며, 타망족(네팔)과 이란에서도 더 낮은 수준으로 나타난다. F1 (P91), F2 (M427), F3 (M481; 이전 F5)는 모두 매우 드물며 스리랑카, 인도, 네팔, 남중국, 태국, 미얀마, 베트남의 지역/소수 민족에 거의 독점적으로 존재한다. F2 (M427)는 동아시아, 인도차이나 반도에서 발견되며, 특히 라후족, 하니족, 이족, 나시족에게서 발견된다.[19]
F xG, H, I, J, K는 현대 인구에서 희귀하며, 특히 남아시아 스리랑카에서 최고 빈도로 나타난다. 또한 동남아시아에서도 오랫동안 존재해 온 것으로 보인다. 술라웨시와 렘바타에서 4~5%의 비율로 보고되었다.
인도네시아 남성을 대상으로 한 연구에 따르면, SNP P14/PF2704(M89와 동일)를 가진 남성은 다음과 같은 비율을 보인다.[18]
4. 6. 하플로그룹 G (M201)
하플로그룹 G (M201)는 약 48,000년 전에 기원했으며, 가장 최근의 공통 조상은 약 26,000년 전 중동에 살았던 것으로 보인다. 이는 신석기 혁명과 함께 유럽으로 확산되었다.이 하플로그룹은 유라시아의 많은 민족 집단에서 발견되며, 특히 코카서스, 이란, 아나톨리아, 레반트 지역에서 흔하게 나타난다. 거의 모든 유럽 국가에서 발견되지만, 가가우지아, 루마니아 남동부, 그리스, 이탈리아, 스페인, 포르투갈, 티롤, 보헤미아에서 가장 흔하게 나타나며, 일부 지중해 섬에서 가장 높은 농도를 보인다. 북유럽에서는 드물게 발견된다.[20][21]
G-M201은 또한 중국 북서부, 인도, 방글라데시, 파키스탄, 스리랑카, 말레이시아, 북아프리카에서도 소수 발견된다.
4. 7. 하플로그룹 H (M69)
하플로그룹 H (M69)는 약 48,000년 전 남부 중앙아시아, 남아시아 또는 서아시아에서 나타난 것으로 추정되며, H1 (M69)과 H3 (Z5857) 형태로 남아시아에 주로 널리 퍼져 있다. 하위 분류군은 이란, 중앙아시아, 중동 전역, 아라비아 반도에서 낮은 빈도로 발견된다.H2 (P96)는 신석기 시대부터 유럽에 존재하며, H1a1 (M82)은 중세 시대에 로마 민족의 이동과 함께 서쪽으로 확산되었다.[72]
4. 8. 하플로그룹 I (M170)
하플로그룹 I (M170, M258)는 주로 유럽과 코카서스에서 발견된다.- 하플로그룹 I1 (M253)은 주로 북유럽에서 발견된다.
- 하플로그룹 I2 (P215)는 주로 발칸 반도, 남동유럽 및 사르데냐에서 발견된다. I2b1 (m223)은 서유럽, 중부 유럽, 북유럽에서 중간 빈도로 발견된다.
4. 9. 하플로그룹 J (M304)
하플로그룹 J는 중동, 코카서스, 남동 유럽에서 주로 발견된다.- 하플로그룹 J1 (M267)는 다게스탄의 북동 코카서스어족 민족과 중동, 에티오피아, 북아프리카의 셈어 사용 민족과 관련이 있으며, 하플로그룹 T와 마찬가지로 지중해 유럽에서도 더 적은 빈도로 발견된다.
- 하플로그룹 J2 (M172)는 주로 셈어 사용 민족, 아나톨리아, 그리스, 발칸 반도, 이탈리아, 이란, 코카서스, 남아시아, 중앙아시아에서 발견된다.
4. 10. 하플로그룹 K (M9)
하플로그룹 K (M9)는 유라시아, 오세아니아, 아메리카 원주민에게 널리 퍼져 있다.K(xLT, K2a, K2b) – 즉, K*, K2c, K2d 또는 K2e –는 주로 멜라네시아, 호주 원주민, 인도, 폴리네시아, 동남아시아 섬 지역에서 발견된다.
4. 11. 하플로그룹 L, T (K1)
L 하플로그룹 (M20)은 남아시아, 중앙아시아, 서남아시아, 지중해 지역에서 발견된다.T 하플로그룹 (M184, M70, M193, M272)은 아프리카의 뿔(주로 쿠시어) 사용 민족) 지역, 남아시아, 중동, 지중해 일부 지역에서 높은 비율로 발견된다. T-M184는 시아카 사람들, 스틸스 사람들, 이집트인, 오만 사람들, 세파르딤 유대인,[22] 이비자(Eivissencs) 사람들, 투부족에서도 상당한 소수에서 발견된다. 또한 지중해 및 남아시아의 다른 지역에서도 낮은 빈도로 발견된다.
4. 12. 하플로그룹 K2 (K-M526)
기저 파라그룹 K2*를 보유한 것으로 보고된 유일한 생존 남성은 호주 원주민이다. 2014년과 2015년에 발표된 주요 연구에 따르면 호주 원주민 남성의 최대 27%가 K2*를 보유하고 있으며, 나머지는 K2의 하위 분류군을 보유하고 있다.[72]4. 13. 하플로그룹 K2a, K2a1, NO & NO1
NO는 북 유라시아에 널리 분포하는 N과 동아시아와 동남아시아에 주로 분포하는 O의 공통 조상이다.- '''N''' 우랄어족과 관련이 있으며, N1a1은 주로 우랄 산맥 서쪽의 핀우고르어파(핀란드인, 에스토니아인, 사미인(랩인종) 등)와 관련이 있고, N1a2는 우랄 산맥 동쪽의 사모예드어파(네네츠인 등)와 관련이 있다. 현대의 중국, 베트남, 조선민주주의인민공화국·대한민국, 일본 등에서도 낮은 빈도로 발견되지만, 고대 인골에서 얻어진 데이터를 고려하면 신석기 시대부터 청동기 시대에 걸쳐 중국 동부(산둥성) 및 북부(허베이성[72], 랴오닝성, 내몽골 자치구)에서 주요 하플로그룹이었던 것으로 보인다.
- '''O'''는 중국 대륙, 한반도, 타이완 섬, 한민족, 조선 민족, 타이인, 미얀마족, 야오족, 셔족, 킨족에 많다. 일본인에게도 15%~20% 정도 발견된다. 시노티베트어족, 먀오야오어족과 관련이 있다.
- O1
- O1a 중국 동남부, 타이완 섬 (특히 타이완 원주민), 해역 동남아시아(필리핀, 말레이시아, 인도네시아, 싱가포르 등)에 많으며, 애드미럴티 제도나 마주로 등에서도 10~30% 정도 발견된다. 오스트로네시아어족과 관련이 있다. 인도차이나 반도, 중국의 기타 지역, 대한민국, 일본, 북아시아, 중앙아시아에서도 낮은 빈도로 발견된다.
- O1b O1b1는 중국 남부, 인도차이나 반도, 인도네시아, 인도 북동부, 니코바르인, 숑펜인, 문다족에 특히 많다. 오스트로아시아어족과의 관련이 상정되고 있지만, 한족이나 일본인에 비교적 많이 발견되는 하위 계통도 현존하고 있다. O1b2는 일본인, 조선 민족에 많고(약 30%), 만주족, 나나이족, 다우르족, 몽골족, 한족 등에도 소수 발견된다. 베트남이나 미크로네시아에서 산발적으로 관찰된 예도 있다. 시모야마혼 유적이나 아오야기카미데라지 유적에서 발견된 야요이 시대의 인골에서 검출된 적이 있다. O1b1과 O1b2의 가장 가까운 공통 조상은 3만 년 이상 전으로 거슬러 올라간다고 추정된다.
- O2 이시카와현가나자와시간포지마치에 있는 이와데 횡혈묘에서 출토된 고분인 한 구의 유골에서 발견된 하플로그룹 O2a2b1a1a1a4a1-CTS5308은 동아시아 전체에서 비교적 많이 발견되는 O-M117의 서브클레이드이지만, O-CTS5308은 특히 티베트인에게 많이 발견되는 타입이다.
4. 14. 하플로그룹 N (M231)
N 하플로그룹(M231)은 동아시아에서 기원하여 북쪽과 서쪽으로 시베리아로 확산되었을 가능성이 있으며, 우랄어족을 사용하는 민족에게서 가장 흔하게 발견되는 그룹이다. 북유라시아에 널리 분포하며, 우랄어족과 관련이 있다. N1a1은 주로 우랄 산맥 서쪽의 핀란드인, 에스토니아인, 사미인(랩인종) 등의 핀우고르어파와 관련이 있고, N1a2는 우랄 산맥 동쪽의 네네츠인 등의 사모예드어파와 관련이 있다. 현대의 중국, 베트남, 대한민국, 조선민주주의인민공화국, 일본 등에서도 낮은 빈도로 발견되지만, 고대 인골에서 얻어진 데이터를 고려하면 신석기 시대부터 청동기 시대에 걸쳐 중국 동부(산둥성) 및 북부(허베이성[72], 랴오닝성, 내몽골 자치구)에서 주요 하플로그룹이었던 것으로 보인다.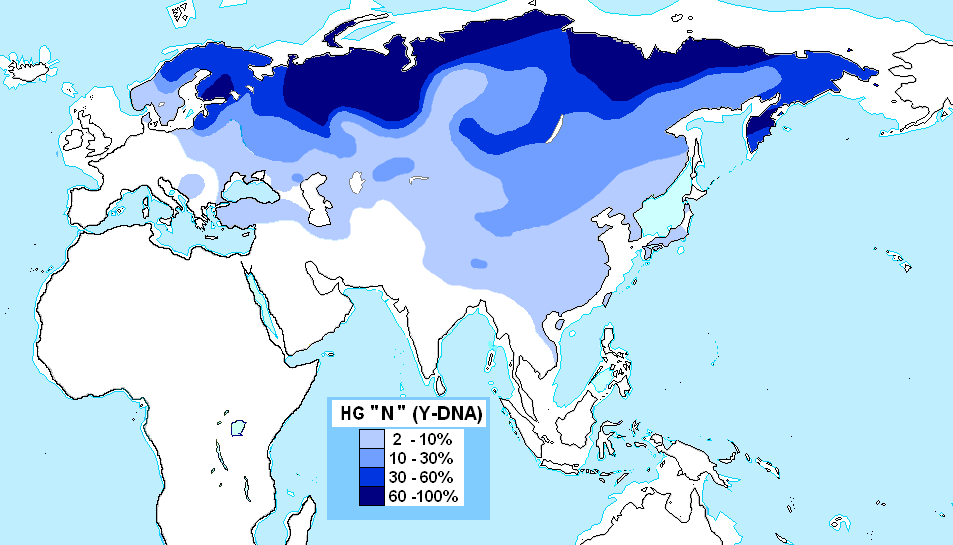
4. 15. 하플로그룹 O (M175)
O 하플로그룹 (M175)은 동아시아와 동남아시아에서 가장 높은 빈도로 발견되며, 오세아니아, 중앙아시아, 남아시아, 인도양의 섬(예: 마다가스카르, 코모로)에서도 낮은 빈도로 발견된다.- O1 하플로그룹 (F265/M1354, CTS2866, F75/M1297, F429/M1415, F465/M1422)
- * O1a 하플로그룹 (M119, CTS31, F589/Page20, L246, L466): 중국 동부, 중부 및 남부, 타이완, 동남아시아에서 발견되며, 특히 오스트로네시아족과 타이-카다이어족에게서 발견된다.
- * O1b 하플로그룹 (P31, M268)
- ** O1b1 하플로그룹 (M95): 일본, 중국, 타이완, 동남아시아, 인도 아대륙에서 발견되며, 특히 오스트로아시아어족과 타이-카다이어족 언어를 사용하는 민족, 말레이족, 인도네시아인에게서 발견된다.
- ** O1b2 하플로그룹 (SRY465, M176): 일본, 한국, 만주, 동남아시아에서 발견된다.
- O2 하플로그룹 (M122): 동아시아, 동남아시아, 오스트로네시아, 폴리네시아 전역에서 발견된다.
4. 16. 하플로그룹 K2b1, M & S
기저 파라그룹 K2b1*의 예는 확인되지 않았다. K2b1의 하위 분류를 가진 남성은 주로 파푸아인, 미크로네시아인, 호주 원주민, 그리고 폴리네시아인 사이에서 발견된다.주요 하위 분류는 다음과 같은 두 개의 주요 하플로그룹이다.
- 하플로그룹 S (B254)는 K2b1a로도 알려져 있으며, 파푸아뉴기니 고지대에서 발견된다.
- 하플로그룹 M (P256)는 K2b1b로도 알려져 있으며, 뉴기니와 멜라네시아에서 발견된다.
4. 17. 하플로그룹 P (K2b2)
하플로그룹 P (P295)는 P1 (P-M45)과 P2 (P-B253)의 두 가지 주요 분기로 나뉜다.[23]P*, P1*, P2는 필리핀 루손섬에서만 발견된다.[23] P*와 P1*은 루손섬의 아에타 (또는 아그타) 사람들 사이에서 많이 발견된다.[24] 반면 P1*은 러시아 극동과 중앙 아시아에서 더 흔하게 발견되며, 동남아시아 본토와 남아시아에서도 낮은 수준으로 발견된다. 이러한 분포는 P*가 K2b에서 동남아시아에서 기원했을 가능성을 시사한다.[24][25]
P1은 하플로그룹 Q (Q-M242)와 하플로그룹 R (R-M207)의 상위 노드이며, 이들은 공통 마커 M45를 공유한다.
4. 18. 하플로그룹 Q (M242)
하플로그룹 P (P295)의 하위 분기인 하플로그룹 Q (Q-M242)는 시베리아와 아메리카 대륙에서 발견된다.[23] 아메리카 원주민에게서 매우 높은 빈도로 나타나며, 케트인 등 시베리아 일부 지역에서도 발견된다.4. 19. 하플로그룹 R (M207)
하플로그룹 R(M207)은 유럽, 서아시아, 중앙아시아, 남아시아 등에서 발견된다. 주요 하위 그룹은 다음과 같다.[73][74]- '''R1a (M420)''' : 동유럽, 북인도, 중앙아시아에 많고, 서 유라시아 전역에서 발견된다. 동유럽 인종, 인도-아프간 인종(인도-아리아인, 이란계 민족)과 관련이 있으며, 인도유럽어족사템어와 관련이 있는 것으로 추정된다.
- '''R1b (M343)''' : 서유럽에 특히 많고, 유럽 전역에서 일반적으로 발견된다. 알프스 인종, 특히 켈트인이나 바스크인에 많다. 인도유럽어족켄툼어와 관련이 있는 것으로 추정된다.
- '''R2 (M124)''' : 남아시아, 코카서스, 중앙아시아, 동유럽에서 발견된다.
5. 한국과 관련된 주요 하플로그룹
한국인중국어의 Y-DNA 하플로그룹은 O1b2가 가장 높은 비율을 차지하고 있으며, 그 외에 D1a2a, C1a1, C2, N 등이 나타난다.
6. 하플로그룹 연대 추정
Y 염색체의 최신 공통 조상(Y-MRCA, 비공식적으로 Y 염색체 아담)은 현재 살아있는 모든 인간이 부계로 자손을 남긴 최신 공통 조상(MRCA)이다.
Y 염색체를 가진 아담은 약 23만 6000년 전에 아프리카에 살았던 것으로 추정된다.
O1b1과 O1b2의 가장 가까운 공통 조상은 3만 년 이상 전으로 거슬러 올라간다.
참조
[1]
논문
A Southeast Asian origin for present-day non-African human Y chromosomes.
[2]
웹사이트
Understanding Haplogroups: How are the haplogroups named?
http://www.familytre[...]
Family Tree DNA
2013-03-31
[3]
논문
Human mutation rate revealed
http://www.nature.co[...]
2017-09-18
[4]
논문
A Southeast Asian origin for present-day non-African human Y chromosomes
2021
[5]
서적
The Prehistory of Language: A Triangulated Y-Chromosome-Based Perspective
https://genlinginter[...]
Genetic–Linguistic Interface
[6]
논문
A recent bottleneck of Y chromosome diversity coincides with a global change in culture
[7]
웹사이트
Something Weird Happened to Men 7,000 Years Ago, And We Finally Know Why
https://www.sciencea[...]
2018-05-31
[8]
웹사이트
Understanding Results: Y-DNA Single Nucleotide Polymorphism (SNP): What is a Y-chromosome DNA (Y-DNA) haplogroup?
http://www.familytre[...]
Family Tree DNA
2013-03-31
[9]
웹사이트
myFTDNA 2.0 User Guide: Y-DNA: What is the Y-DNA – Matches page?
http://www.familytre[...]
Family Tree DNA
2013-03-31
[10]
웹사이트
Understanding Results: Y-DNA Single Nucleotide Polymorphism (SNP): How are haplogroups and their subclades named?
http://www.familytre[...]
Family Tree DNA
2013-03-31
[11]
웹사이트
New Y DNA Haplogroup Naming Convention
https://dna-explaine[...]
2024-08-31
[12]
웹사이트
ISOGG 2015 Y-DNA Haplogroup Tree Trunk
http://www.isogg.org[...]
[13]
논문
Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations
[14]
논문
New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree
[15]
논문
A recent bottleneck of Y chromosome diversity coincides with a global change in culture
2015-04
[16]
논문
A Rare Deep-Rooting D0 African Y-chromosomal Haplogroup and its Implications for the Expansion of Modern Humans Out of Africa
2019-06
[17]
논문
Y chromosome diversity, human expansion, drift, and cultural evolution
2009-12-01
[18]
논문
Isolation, contact and social behavior shaped genetic diversity in West Timor
[19]
문서
This was, for instance, the case with the original subclade F3 (M96), which has since been renamed [[Haplogroup H (Y-DNA)#Haplogroup H2 (P96)|Haplogroup H2]].
[20]
논문
Different genetic components in the Norwegian population revealed by the analysis of mtDNA and Y chromosome polymorphisms
[21]
논문
Y-chromosome diversity in Sweden – A long-time perspective
[22]
논문
Phylogeographic analysis of paternal lineages in NE Portuguese Jewish communities
[23]
웹사이트
ISOGG, 2016, ''Y-DNA Haplogroup P and its Subclades – 2016''
http://isogg.org/tre[...]
2016-06-20
[24]
논문
Isolation, contact and social behavior shaped genetic diversity in West Timor
2014-07-31
[25]
논문
Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia
2015
[26]
논문
Mitochondrial Population Genomics Supports a Single Pre-Clovis Origin with a Coastal Route for the Peopling of the Americas
http://www.familytre[...]
2013-05-22
[27]
논문
High-Resolution SNPs and Microsatellite Haplotypes Point to a Single, Recent Entry of Native American Y Chromosomes into the Americas
[28]
웹사이트
Y-DNA Haplogroup Tree 2010
http://www.isogg.org[...]
International Society of Genetic Genealogy
2010-07-01
[29]
논문
A novel subgroup Q5 of human Y-chromosomal haplogroup Q in India
[30]
논문
The Y-Chromosome Tree Bursts into Leaf: 13,000 High-Confidence SNPs Covering the Majority of Known Clades
[31]
논문
Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge
[32]
논문
Genetic evidence supports demic diffusion of Han culture
2004-09
[33]
논문
The Eurasian heartland: a continental perspective on Y-chromosome diversity
2001-08
[34]
논문
Y-chromosome evidence for differing ancient demographic histories in the Americas
2003-09
[35]
논문
Large-scale recent expansion of European patrilineages shown by population resequencing
[36]
웹사이트
YFull YTree
https://www.yfull.co[...]
YFull
2017-09-15
[37]
논문
The Divergence of Neandertal and Modern Human Y Chromosomes
[38]
웹사이트
The father of all men is 340,000 years old
https://www.newscien[...]
[39]
논문
A recent bottleneck of Y chromosome diversity coincides with a global change in culture
2015-04
[40]
논문
A Rare Deep-Rooting D0 African Y-chromosomal Haplogroup and its Implications for the Expansion of Modern Humans Out of Africa
2019-06
[41]
논문
Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area
[42]
논문
Origin, diffusion, and differentiation of Y-chromosome haplogroups E and J: inferences on the neolithization of Europe and later migratory events in the Mediterranean area
2004-05
[43]
논문
Genetic Evidence of an East Asian Origin and Paleolithic Northward Migration of Y-chromosome Haplogroup N
[44]
논문
Phylogeographic Refinement and Large Scale Genotyping of Human y Chromosome Haplogroup e Provide New Insights into the Dispersal of Early Pastoralists in the African Continent
[45]
논문
Large-scale recent expansion of European patrilineages shown by population resequencing
[46]
간행물
New Phylogenetic Relationships for Y-chromosome Haplogroup I: Reappraising its Phylogeography and Prehistory
P. Mellars, K. Boyle, O. Bar-Yosef and C. Stringer (eds.), Rethinking the Human Evolution
2007
[47]
논문
The Indian origin of paternal haplogroup R1a1* substantiates the autochthonous origin of Brahmins and the caste system
[48]
웹사이트
ftDNA
https://www.familytr[...]
[49]
문서
Myres2010
[50]
논문
Upper Palaeolithic genomes reveal deep roots of modern Eurasians
[51]
논문
Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe
http://www.familytre[...]
2016-05-04
[52]
웹사이트
Mesolithic Western Eurasian DNA
http://www.ancestral[...]
2016-05-04
[53]
서적
The Evatt Clan: A Worldwide Historical Review of the Evatt Family Surname
https://books.google[...]
2013-11-01
[54]
논문
Wave-of-Advance Models of the Diffusion of the y Chromosome Haplogroup R1b1b2 in Europe
[55]
논문
Tracing Past Human Male Movements in Northern/Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12
[56]
논문
Separating the post-Glacial coancestry of European and Asian y chromosomes within haplogroup R1a
[57]
논문
Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome
[58]
웹사이트
'Y-DNA Haplogroup Tree 2015'
http://www.isogg.org[...]
International Society of Genetic Genealogy (ISOGG; 2015)
[59]
문서
Haplogroup A0-T is also known as A-L1085 (and previously as A0'1'2'3'4).
[60]
문서
Haplogroup A1 is also known as A1'2'3'4.
[61]
문서
F-Y27277, sometimes known as F2'4, is both the parent clade of F2 and F4 and a child of F-M89.
[62]
문서
Haplogroup LT (L298/P326) is also known as Haplogroup K1.
[63]
문서
Between 2002 and 2008, [[Haplogroup T-M184]] was known as "Haplogroup K2". That name has since been re-assigned to [[Haplogroup K2|K-M526]], the sibling of Haplogroup LT.
[64]
문서
Haplogroup K2b (M1221/P331/PF5911) is also known as Haplogroup MPS.
[65]
문서
Haplogroup K2b1 (P397/P399) is also known as Haplogroup MS, but has a broader and more complex internal structure.
[66]
문서
Haplogroup P (P295) is also klnown as K2b2.
[67]
문서
K-M2313*, which as yet has no phylogenetic name, has been documented in two living individuals, who have ethnic ties to India and South East Asia. In addition, K-Y28299, which appears to be a primary branch of K-M2313, has been found in three living individuals from India.
[68]
문서
Haplogroup S, as of 2017, is also known as K2b1a. (Previously the name Haplogroup S was assigned to K2b1a4.)
[69]
문서
Haplogroup M, as of 2017, is also known as K2b1b. (Previously the name Haplogroup M was assigned to K2b1d.)
[70]
간행물
The East Asian linguistic phylum: A reconstruction based on language and genes
2018
[71]
문서
F36の分岐指標はFTDNAにあるが、Y Fullにはない。
[72]
논문
Genetic diversity of two Neolithic populations provides evidence of farming expansions in North China.
2017
[73]
서적
新日本人の起源
勉誠出版
2009
[74]
서적
DNA・考古・言語の学際研究が示す 新・日本列島史
勉誠出版
2009
[75]
논문
A Southeast Asian origin for present-day non-African human Y chromosomes
2021-02
[76]
서적
人類史マップ サピエンス誕生・危機・拡散の全記録
[77]
서적
人類史マップ サピエンス誕生・危機・拡散の全記録
[78]
웹인용
PhyloTree.org
http://www.phylotree[...]
[79]
웹인용
PhyloTree.org
http://www.phylotree[...]
[80]
웹인용
PhyloTree.org
http://www.phylotree[...]
[81]
서적
人類史マップ サピエンス誕生・危機・拡散の全記録
[82]
웹인용
ISOGG 2015 Y-DNA Haplogroup Tree Trunk
http://www.isogg.org[...]
[83]
논문
Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe
http://www.cell.com/[...]
2004
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com