식세포
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
식세포는 세균, 기생충, 죽은 세포 등 이물질을 섭취하는 과정을 의미한다. 1882년 러시아의 생물학자 일리야 메치니코프가 불가사리 유생 연구를 통해 처음 인지했으며, 1908년에는 식세포와 식작용에 대한 공로를 인정받아 노벨 생리학·의학상을 수상했다. 식작용은 식세포 표면의 수용체와 외부 물질의 결합으로 시작되며, 세균은 파고솜에 갇히고 리소좀과 융합하여 파고리소좀을 형성하여 살해된다. 식세포는 전문 식세포와 비전문 식세포로 구분되며, 종류에 따라 활성화, 이동, 살균 방법 등이 다르다. 병원체는 식세포의 공격을 회피하거나, 식세포 안에서 생존하는 등 다양한 방법으로 면역계를 회피한다. 식세포는 숙주 조직을 손상시키기도 하며, 진화 초기에 단세포 진핵생물에서 나타난 것으로 추정된다.
더 읽어볼만한 페이지
- 식세포 - 세포 매개 면역
세포 매개 면역은 항체의 도움 없이 T 세포, 자연 살해 세포(NK 세포), 대식세포 등의 세포들이 직접 항원을 제거하는 면역 반응으로, 세포독성 T 세포는 감염 세포나 암세포를 파괴하고, NK 세포는 비자기 세포를 파괴하며, 대식세포는 식균 작용을 통해 병원체를 제거하고, 사이토카인을 분비하여 면역 반응을 조절하며, 바이러스 감염, 암, 이식 거부 반응 등에 관여한다. - 식세포 - 호중구
호중구는 혈액 내에서 가장 풍부한 백혈구로서 세균 감염에 대한 신속한 면역 반응에 중요한 역할을 하며, 식세포 작용, 탈과립, 호중구 세포외덫 형성 등의 항미생물 작용을 수행하고 염증 반응을 조절하며 사이토카인을 분비한다. - 면역계 - 알레르기
알레르기는 면역 체계가 무해한 물질에 과도하게 반응하여 아토피 피부염, 비염, 천식, 두드러기 등 다양한 증상을 일으키는 질환으로, Gell과 Coombs 분류법에 따라 여러 유형으로 나뉘며, 심각한 경우 아나필락시스를 유발할 수 있고, 유전적, 환경적 요인이 복합적으로 작용하여 발생하며, 알레르겐 회피, 약물, 면역요법 등으로 치료한다. - 면역계 - 면역억제
면역억제는 면역계의 활동을 감소시키는 현상으로, 의도적으로는 장기 이식 거부 반응 방지나 자가면역 질환 치료를 위해 면역억제제를 사용하고, 비의도적으로는 질병으로 인해 발생하며 면역결핍질환을 유발할 수 있다. - 알찬 글 - 동성애
동성애는 동성 간의 성적 매력, 행위, 애정을 의미하는 용어로, LGBT와 같은 포괄적인 용어가 사용되며 사회적 인식 변화와 함께 동성애자 권리 보호 노력이 진행 중이나, 원인은 명확히 밝혀지지 않았다. - 알찬 글 - 올림픽
올림픽은 고대 그리스에서 시작되어 근대에 부활한 국제 스포츠 경기 대회로, 하계 및 동계 올림픽, 패럴림픽, 청소년 올림픽 등으로 나뉘어 국제올림픽위원회(IOC)가 총괄하지만, 개최 비용, 정치적 이용, 도핑, 인권 문제 등의 논란이 있다.

2. 역사

러시아의 생물학자 엘리 메치니코프(1845–1916)는 미생물 감염을 방어하는 데 특화된 세포가 존재한다는 사실을 처음으로 인지했다.[156][13] 1882년, 그는 불가사리 유생에 있는 운동성 세포들을 연구하며 이 세포들이 동물의 면역에 중요할 것이라고 생각했다. 이를 확인하기 위해 그는 유생에 귤나무의 작은 가시를 삽입했고, 몇 시간 뒤 가시 주변으로 운동성 세포들이 모여드는 것을 관찰했다.[156][13] 메치니코프는 빈에서 카를 프리드리히 빌헬름 클라우스에게 이 발견을 알렸고, 클라우스는 이 세포에 '식세포(phagocyte)'라는 이름을 제안했다. 이는 '먹다'라는 뜻의 그리스어 phagein|파게인grc과 '세포'를 의미하는 kutos|퀴토스grc에서 유래한 이름이다.[141][157][14][15]
약 1년 후, 메치니코프는 물벼룩을 연구하던 중, 물벼룩을 공격한 진균의 포자가 식세포에 의해 파괴되는 것을 관찰했다. 그는 이 관찰을 포유류의 백혈구로 확장하여, 식세포가 탄저균(''Bacillus anthracis'')을 삼켜 죽일 수 있다는 것을 발견하고 이 과정을 '식작용(phagocytosis)'이라고 명명했다.[158][16] 메치니코프는 식세포가 외부 침입 유기체에 대한 인체의 일차 방어선이라고 주장했다.[156][13]
한편, 1903년 암로스 라이트는 식작용이 특정 항체에 의해 강화된다는 사실을 발견하고, 이 항체에 '옵소닌(opsonin)'이라는 이름을 붙였다. 이는 '샐러드 드레싱'이나 '렐리시'를 의미하는 그리스어 단어 ὄψον|옵손grc에서 유래했다.[159][17]
메치니코프는 식세포와 식작용 연구에 대한 공로를 인정받아 파울 에를리히와 함께 1908년 노벨 생리학·의학상을 수상했다.[146][4]
이러한 발견들은 20세기 초에 서서히 받아들여졌으나, 식세포와 다른 면역계 요소들 간의 복잡한 상호작용은 1980년대에 이르러서야 완전히 이해되기 시작했다.[160][18]
3. 식작용

식작용은 세균, 기생충, 세포자멸사로 죽은 숙주 세포, 세포의 잔해와 같은 입자들을 세포가 섭취하는 과정이다.[161][20] 이 과정은 여러 분자적 단계들이 연쇄적으로 일어나며 진행된다.[162][21][22]
식작용은 외부 물질이나 세균 등이 식세포 표면에 있는 수용체 분자와 결합하면서 시작된다.[19] 결합이 일어나면 식세포는 세포막을 늘려 외부 물질 주변을 둘러싸며 안으로 삼킨다. 사람의 호중구가 세균 하나를 식작용하는 데에는 평균적으로 약 9분이 걸린다.[163][23] 식세포 안으로 들어온 외부 물질은 파고솜이라는 소포 안에 갇히게 된다. 약 1분 안에 이 파고솜은 세포 내 소화기관인 리소좀이나 과립과 융합하여 파고리소좀을 형성한다.[164][24] 파고리소좀 안에서 외부 물질은 강력한 살균 및 소화 효소 등에 의해 분해되며,[164][24] 세균의 경우 보통 몇 분 안에 사멸한다.[163][23]
수지상세포와 대식세포의 식작용은 호중구보다 느려서, 때로는 몇 시간이 걸리기도 한다.[165][25] 특히 대식세포는 느리지만 많은 양의 물질을 탐식하며, 소화되지 않은 일부 물질을 조직으로 다시 방출하기도 한다. 이렇게 방출된 파편들은 혈액 속의 다른 식세포들을 문제 부위로 불러 모으는 신호 역할을 한다.[165][25] 식세포의 식욕은 매우 왕성해서, 과학자들이 철가루를 대식세포에게 먹인 뒤 자석을 이용해 다른 세포들로부터 분리하는 실험을 하기도 했다.[165][26]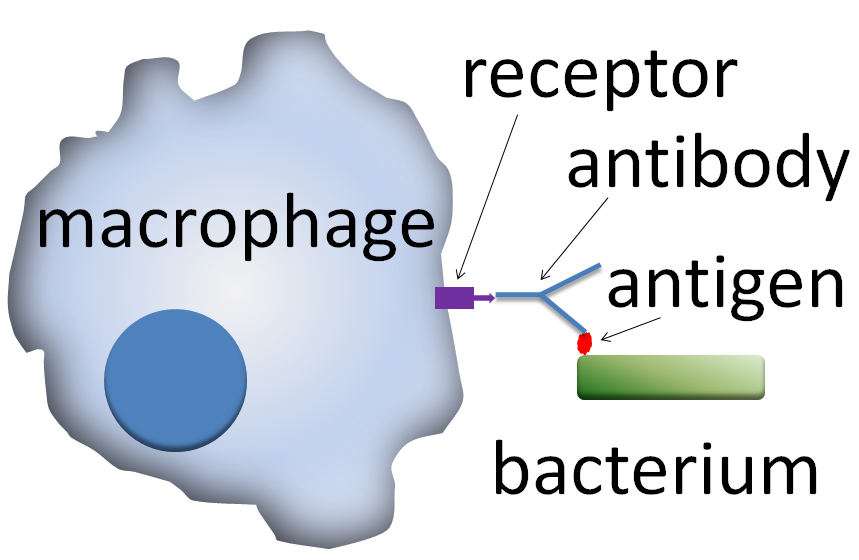
식세포는 표면에 다양한 종류의 수용체를 가지고 있어 외부 물질과 효과적으로 결합한다.[142][12] 주요 수용체는 다음과 같다.
4. 살균 방법
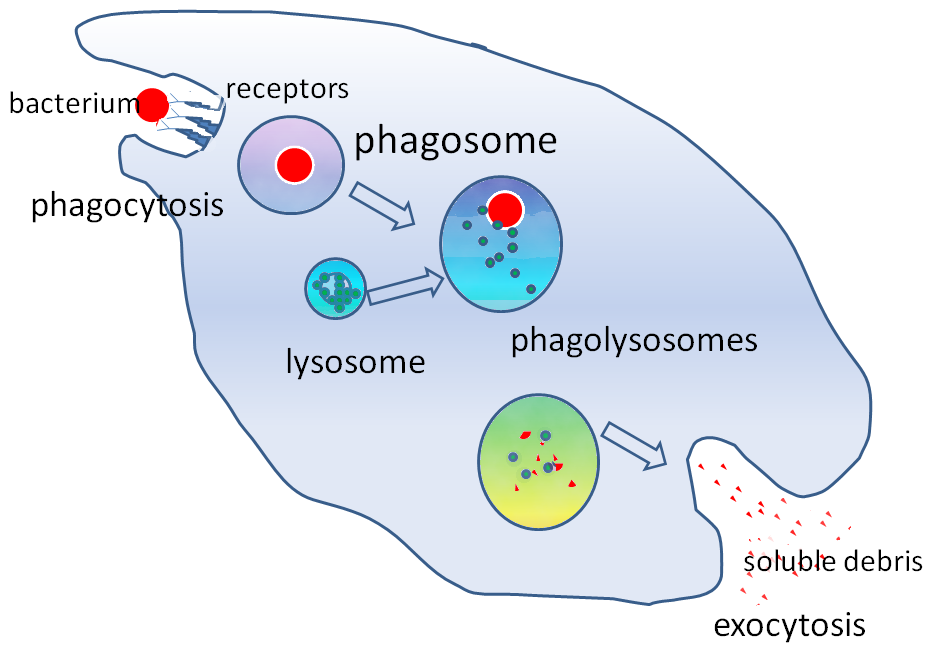
미생물을 죽이는 것은 식세포의 중요한 작용으로, 식세포 안쪽(세포내 살균, intracellular killing)과 바깥쪽(세포외 살균, extracellular killing) 모두에서 일어난다.[167][29]
=== 세포 내 살균 ===
식세포가 세균이나 다른 물질을 삼키면 산소 소비량이 급격히 증가하는데, 이를 호흡폭발(respiratory burst)이라고 한다. 호흡폭발 과정에서 식세포는 강력한 항미생물 활성을 가진 활성 산소(영어: Reactive Oxygen Specieseng, ROS)를 포함하는 물질들을 생산한다.[168][30] 이러한 산소 화합물들은 침입한 미생물뿐만 아니라 식세포 자신에게도 해로울 수 있어, 평소에는 세포 내 특정 공간에 격리되어 보관된다.[144][2] 활성 산소를 이용해 미생물을 죽이는 방식을 산소 의존성 세포내 살균(oxygen-dependent intracellular killing)이라고 하며, 두 가지 주요 유형이 있다.[144][2]
첫 번째 유형은 초과산화물(superoxide, O2-)을 산소에 의존하여 생산하는 것이다.[142][12] 초과산화물은 산소가 풍부한 환경에서 세균을 효과적으로 죽일 수 있는 물질이다.[169] 이 초과산화물은 초과산화물 불균등화효소(superoxide dismutase)라는 효소에 의해 과산화 수소(H2O2)와 산소(O2) 분자로 전환된다. 또한, 초과산화물은 과산화 수소와 반응하여 매우 강력한 살균력을 지닌 하이드록실 라디칼(hydroxyl radical, •OH)을 생성하여 미생물 제거를 돕는다.[142][12]
두 번째 유형은 주로 호중구의 과립에 존재하는 골수세포형과산화효소(myeloperoxidase, MPO)를 이용하는 방식이다.[170][32] 식세포가 미생물을 삼켜 형성된 파고솜(phagosome)이 세포 내 과립과 융합하면, MPO가 파고리소좀(phagolysosome) 안으로 방출된다. 이 효소는 과산화 수소와 염소 이온(Cl-)을 이용하여 차아염소산염(hypochlorite, OCl-)을 생성한다. 차아염소산염은 가정용 표백제의 주성분으로, 세균에게 매우 치명적인 독성을 나타낸다.[142][12] MPO는 헴(heme) 색소를 포함하고 있어, 고름이나 감염된 객담(가래)과 같이 호중구가 많이 모인 분비물이 녹색을 띠게 하는 원인이 된다.[171][33]
식세포는 산소를 사용하지 않는 산소 비의존성 세포내 살균(oxygen-independent intracellular killing) 방법으로도 미생물을 죽일 수 있지만, 이는 산소 의존적인 방법에 비해 효과는 다소 떨어진다. 여기에는 네 가지 주요 방식이 있다.
# 세균의 세포막에 손상을 주는 전하를 띈 단백질을 이용한다.
# 리소좀(lysosome)에 포함된 효소(예: 리소자임)를 이용하여 세균의 세포벽을 분해한다.
# 호중구 과립에 존재하는 락토페린(lactoferrin)을 이용하여 세균 생존에 필수적인 철 이온을 빼앗는다.[172][34]
# 단백질분해효소(protease)와 가수분해효소(hydrolase)를 이용하여 파괴된 세균의 단백질 등을 최종적으로 소화한다.[173][35]
=== 세포 외 살균 ===
식세포, 특히 대식세포는 세포 밖에서도 미생물을 죽일 수 있다. 인터페론 감마(Interferon-gamma, IFN-γ)는 대식세포를 활성화시켜 일산화 질소(Nitric oxide, NO)를 생산하도록 유도한다. 인터페론 감마는 보조 T세포(CD4+ T 세포), 세포독성 T세포(CD8+ T 세포), 자연 살해 세포(NK 세포), 단핵구, 다른 대식세포, 수지상세포 등 다양한 면역 세포에서 분비될 수 있다.[174][36] 대식세포에서 방출된 일산화 질소는 그 자체의 독성으로 주변에 있는 미생물을 효과적으로 제거한다.[142][12]
또한, 활성화된 대식세포는 종양괴사인자(Tumor necrosis factor, TNF)를 생산하고 분비한다. TNF는 사이토카인의 일종으로, 암세포나 바이러스에 감염된 세포를 직접 죽이는 역할도 하며, 다른 면역 세포들을 활성화시켜 면역 반응을 증폭시키는 중요한 신호 분자로 작용한다.[175][37][176][38]
=== 살균 능력의 결함 ===
드물지만 선천적으로 식세포의 살균 능력에 문제가 있는 질병도 존재한다. 예를 들어, 만성 육아종병(Chronic granulomatous disease)은 산소 의존성 살균 과정에 필요한 효소 등에 이상이 생겨, 세균 감염이 반복적으로 발생하는 유전 질환이다.[177][39] 체디아크-히가시 증후군(Chédiak–Higashi syndrome) 역시 식세포가 삼킨 미생물을 제대로 죽이지 못하는 희귀 질환 중 하나이다.[178][40]
5. 세포자살에서의 역할
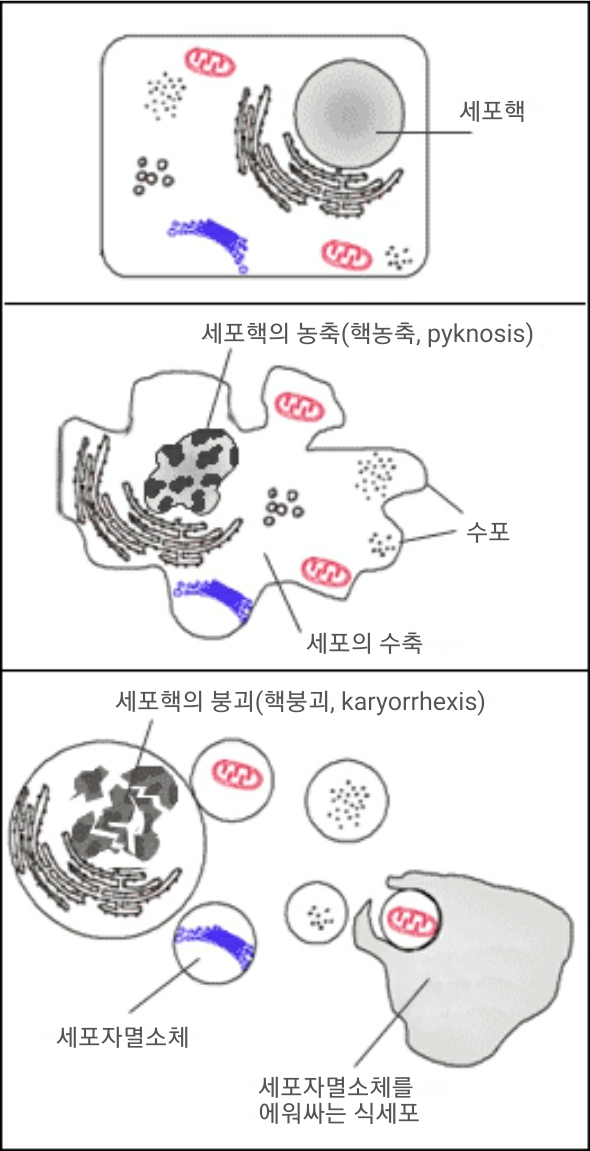
동물에서 세포들은 수명이 다하면 꾸준히 죽는다. 세포분열과 죽는 세포 간에 균형이 맞춰져 있으므로, 성인에서 세포의 숫자는 비교적 잘 유지된다.[152] 세포가 죽는 데에는 괴사와 세포자살이라는 두 가지 서로 다른 방법이 있다. 괴사는 종종 질병이나 외상에 의해 일어나지만 세포자살(또는 세포예정사, programmed cell death)은 세포의 정상적인 기능이다. 몸은 죽었거나 죽어가는 중인 자신의 세포를 매일 수백만 개씩 제거해야 하며, 식세포는 이 제거 과정에 핵심적인 역할을 한다.[182]
세포자살의 마지막 단계를 거치는 중인 죽어가는 세포들[183]은 식세포를 유인하기 위해 포스파티딜세린과 같은 분자들을 표면에 발현한다.[184] 포스파티딜세린은 세포막의 세포질 쪽 표면에서 정상적으로 발견되지만 세포자살 과정이 일어날 때는 스크렘블레이스라는 단백질에 의해 세포막의 세포 바깥쪽 표면으로 위치가 바뀐다.[185][186] 이 분자들은 대식세포와 같이 적절한 수용체를 가진 세포에 의한 식작용이 일어날 세포를 표시하는 역할을 한다.[187] 식세포에 의해 죽는 세포를 제거하는 과정은 염증 반응을 유발하지 않는 평화로운 방식으로 일어나며, 식세포의 중요한 기능 중 하나이다.[188]
6. 다른 세포들과의 상호작용
식세포는 특정 기관에 고정되어 있지 않고, 몸 전체를 자유롭게 이동하며 다른 식세포나 면역계의 다른 세포들과 끊임없이 상호작용한다.[52] 이들은 사이토카인이라는 화학 신호 물질을 분비하여 다른 세포들과 소통하는데, 이를 통해 감염이 발생한 곳으로 다른 식세포들을 불러 모으거나, 아직 활동하지 않는 휴면 상태의 림프구를 자극하여 면역 반응을 개시하고 조절한다.[189][52]
식세포는 인간을 포함한 동물이 태어날 때부터 가지고 있는 선천면역 시스템의 핵심적인 역할을 담당한다. 선천면역은 매우 효과적이지만, 침입한 대상의 종류를 세세하게 구별하지는 못하는 비특이적 방어 방식이다.[190][53] 이와 대조적으로, 유악류(턱이 있는 척추동물)에서 발달한 적응면역은 특정 병원체에 맞춰 고도로 전문화된 방어 체계로, 주로 림프구가 그 역할을 수행한다.[190][53] 림프구는 항체라는 특수한 단백질을 만들어 침입자를 직접 무력화하거나 다른 면역 세포가 파괴하기 쉽도록 표지하는 역할을 한다.[191][54]
식세포, 특히 대식세포와 수지상세포는 선천면역과 적응면역 사이를 연결하는 중요한 다리 역할을 수행한다. 이들은 항원제시라는 과정을 통해 자신이 처리한 병원체의 정보를 림프구에게 전달함으로써, 림프구가 특정 항원에 효과적으로 반응하여 항체를 생산하도록 돕는다.[192][55]
또한 식세포는 다양한 사이토카인을 통해 다른 세포들과 상호작용하며 면역 반응을 조절한다. 예를 들어, 인터페론 감마(IFN-γ)는 대식세포를 활성화시켜 독성 물질인 일산화 질소(NO)를 생산하게 만든다. 이 일산화 질소는 대식세포 주변의 미생물을 직접 살상하는 효과를 가진다.[174][142][36][12] 활성화된 대식세포는 종양괴사인자(TNF)라는 또 다른 중요한 사이토카인을 생산하고 분비하는데,[175][37] 이는 암 세포나 바이러스에 감염된 세포를 직접 파괴하고 면역계의 다른 세포들을 활성화시키는 등 다방면에 걸쳐 면역 반응을 조절하는 역할을 수행한다.[176][38]
6. 1. 항원제시

항원제시는 일부 식세포가 탐식한 물질의 일부를 자신의 세포 표면으로 옮겨 면역계의 다른 세포들에게 보여주는 과정이다.[193][56] 이 역할을 전문적으로 수행하는 세포를 전문 항원제시세포라고 하며, 대표적으로 대식세포와 수지상세포가 있다.[194][57] B 세포 역시 항원 제시 기능을 수행한다. 식세포, 특히 수지상 세포와 대식세포는 항원 제시라는 중요한 과정을 통해 림프구가 항체를 생성하도록 자극한다.[55]
식세포가 외부 단백질과 같은 항원을 탐식하면, 세포 내부에서 이를 잘게 부순 펩타이드 조각으로 만든다. 이 펩타이드 조각들은 세포 안에 있는 주조직 적합성 복합체(MHC) 분자라는 당단백질(특히 MHC 클래스 II 분자)와 결합한다.[155][11] MHC 분자는 이 펩타이드 조각을 식세포의 표면으로 운반하여, 림프구, 특히 헬퍼 T 세포(CD4+)에게 항원으로서 제시한다.[155][11] 헬퍼 T 세포는 제시된 항원을 인식하여 세포성 면역 반응을 활성화시키는 데 중요한 역할을 한다.
성숙한 대식세포는 주로 감염이 일어난 부위에 머무르지만, 수지상세포는 다르다. 수지상세포는 탐식한 항원 정보를 가지고 몸속 림프절로 이동할 수 있다.[195][58] 림프절에는 수많은 림프구가 모여 있어, 수지상세포가 가져온 항원 정보에 효과적으로 반응하여 면역 반응을 더욱 강화시킬 수 있다.[196][59]
한편, 수지상세포가 외부 항원이 아닌 우리 몸 자신의 구성 성분(자가 항원)을 림프구에게 제시하는 경우도 있다. 이때는 면역 반응을 일으키는 대신 오히려 해당 림프구를 파괴하거나 활동하지 못하게 만든다. 이는 우리 몸이 스스로를 공격하는 자가 면역 질환을 예방하는 데 필수적인 과정이며, 이를 면역관용(immune tolerance) 또는 내성이라고 부른다.[197][60]
6. 2. 면역관용
수지상세포는 우리 몸이 스스로를 공격하는 것을 막는 면역관용을 촉진하는 중요한 역할을 한다.[198][61] 면역관용은 크게 두 단계로 나눌 수 있다.첫 번째 단계는 가슴샘에서 일어나는 중추관용이다. 이 과정에서 수지상세포는 MHC 분자에 붙인 자가 항원을 제시하는데, 만약 T세포의 T세포 수용체가 이 자가 항원에 너무 강하게 결합하면 해당 T세포는 제거(사멸)된다. 이를 통해 우리 몸에 강하게 반응하는 T세포를 미리 걸러낸다.
두 번째 단계는 말초관용이다. 여러 이유로, 특히 가슴샘에서 일부 자가 항원의 발현이 부족하여 소수의 자가반응성 T세포가 중추관용 단계를 통과하여 가슴샘 밖, 즉 말초로 나갈 수 있다. 말초에서는 조절 T세포와 같은 다른 종류의 T세포가 이러한 자가반응성 T세포의 활동을 억제하거나 조절하여 면역 반응이 과도하게 일어나지 않도록 막는다.[199][62]
만약 이러한 면역관용 과정이 제대로 작동하지 않으면, 면역계가 자신의 조직이나 세포를 공격하는 자가 면역 질환이 발생할 수 있다.[200][63]
7. 전문 식세포

사람과 다른 유악류의 식세포는 식작용 관여 시의 효율에 따라 '전문'(professional) 식세포와 '비전문'(non-professional) 식세포로 나누어진다.[149] 전문 식세포에는 단핵구, 대식세포, 호중구, 조직 수지상세포, 비만세포가 있다.[146] 1L의 사람 혈액에는 대략 60억 개의 식세포가 포함되어 있다.[145]
7. 1. 활성화
모든 식세포, 특히 대식세포는 항상 특정 준비 상태에 있다. 대식세포는 대부분의 조직에서 비교적 활동하지 않는 상태로 존재하며 천천히 증식한다. 이 준-휴식 상태에서는 주로 몸의 죽은 세포나 감염과 관련 없는 물질들을 제거하며, 항원 제시에는 거의 참여하지 않는다.그러나 감염이 발생하면, 대식세포는 인터페론 감마와 같은 화학적 신호를 받아 활성화된다. 이 신호는 제2형 주조직 적합성 복합체(MHC class II)의 생산을 늘려 항원을 제시할 준비를 갖추게 한다. 활성화된 대식세포는 항원 제시 능력과 병원체를 죽이는 능력이 향상된다. 만약 침입한 병원체로부터 직접적인 신호를 받으면, 대식세포는 '과활성화' 상태가 된다. 이 상태에서는 증식을 멈추고 오직 병원체 제거에만 집중한다. 대식세포의 크기가 커지고 식작용 속도도 빨라지며, 일부는 침입한 원생동물을 통째로 삼킬 수 있을 정도로 커지기도 한다.[201][64]
혈액 속의 호중구는 평소 비활성 상태로 혈류를 따라 빠르게 이동한다. 염증이 생긴 부위의 대식세포로부터 신호를 받으면, 호중구는 이동 속도를 늦추고 혈관을 빠져나와 조직으로 이동한다. 조직에 도착한 호중구는 사이토카인에 의해 활성화되어 병원체를 제거할 준비를 마친 상태로 감염 현장에 도달한다.[202][65]
7. 2. 이동

감염이 일어나면, 감염된 장소로 식세포를 끌어 모으기 위한 화학적인 'SOS' 신호가 발생한다.[203] 이 화학 신호는 침투한 세균에서 나온 단백질, 혈액 응고 과정에서 만들어지는 펩타이드, 보체계의 생산물, 또는 감염된 곳 근처 조직에 있던 대식세포가 방출하는 사이토카인 등을 포함할 수 있다.[142] 특히 사이토카인은 혈액으로부터 호중구와 단핵구를 감염 부위로 불러 모으는 역할을 한다.[153]
식세포가 감염 장소에 도착하려면 혈류를 떠나 감염된 조직으로 들어가야 한다. 감염 신호는 혈관 안쪽 벽을 덮고 있는 내피세포가 셀렉틴이라는 단백질을 만들도록 유도한다. 이동 중인 호중구는 이 셀렉틴에 달라붙어 혈관 벽을 따라 이동 속도를 늦춘다. 다른 신호들은 내피세포 사이의 연결(접합부)을 느슨하게 만들어, 식세포가 혈관 벽 틈새를 비집고 빠져나갈 수 있게 한다. 이렇게 혈관 밖으로 나간 식세포는 주화성이라는 과정을 통해 사이토카인 농도가 높은 감염 장소를 향해 이동한다.[142] 호중구는 상피세포로 덮인 조직을 가로질러 이동하는데, 이 과정은 감염과 싸우는 데 매우 중요하지만, 때로는 이동 과정 자체가 염증 반응을 일으켜 질병과 유사한 증상을 유발하기도 한다.[204] 감염이 발생하면 수백만 개의 호중구가 혈액에서 동원되어 감염 부위로 이동하지만, 이들은 수명이 짧아 며칠 안에 사멸한다.[205]
7. 3. 단핵구
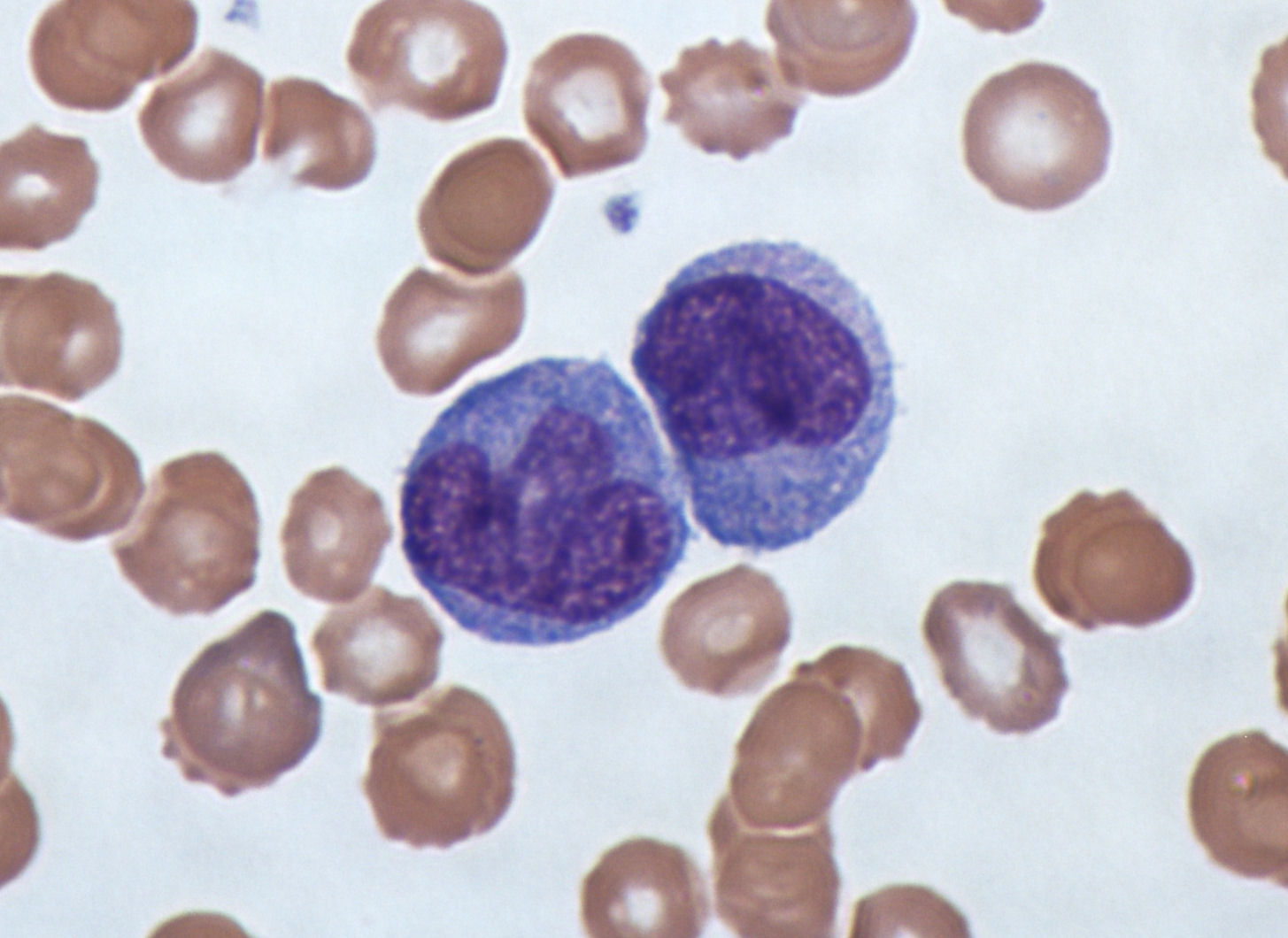
단핵구는 골수에서 발생하여 혈액에서 성숙한다. 성숙한 단핵구는 크고 부드러운, 엽으로 나누어진 핵을 가지며 과립을 포함한 풍부한 세포질로 이루어져 있다. 단핵구는 이물질이나 위험한 물질을 섭취하여 면역계의 다른 세포에게 항원으로 제시한다. 단핵구는 순환하는(circulating) 그룹과 가장자리(marginal) 그룹의 두 그룹을 형성하며, 대략 70% 정도는 가장자리 그룹이다. 대다수 단핵구는 20-40시간이 지나면 혈류를 떠나 조직이나 장기로 이동하고, 그곳에서 받은 신호에 따라 대식세포[206]나 수지상세포로 변한다.[207] 사람 혈액 1L에는 약 5억 개의 단핵구가 포함되어 있다.[145]
7. 4. 대식세포
성숙한 대식세포(macrophage)는 멀리 이동하지 않고 주로 외부 환경에 노출된 신체 부위를 방어하는 역할을 수행한다. 위치한 조직에서 받는 신호에 따라 죽은 세포나 잔해를 처리하는 청소부 역할을 하거나(식작용), 항원제시세포로서 면역계의 다른 세포에게 외부 침입 정보를 전달하며, 때로는 강력한 살균 세포로 변모하여 병원체를 직접 공격한다.[72][208] 대식세포는 혈액 속 단핵구가 조직으로 이동하여 분화하거나, 골수의 과립구 줄기세포로부터 유래하며, 이미 조직에 존재하는 대식세포가 세포분열을 통해 증식하여 형성되기도 한다.[73][209] 사람의 대식세포 크기는 직경 약 21마이크로미터이다.[74][210]
대식세포는 호중구와 달리 세포 내부에 뚜렷한 과립을 가지지 않지만, 소화 효소가 풍부한 리소좀을 다수 포함하고 있다.[211] 이들은 우리 몸의 거의 모든 조직과 장기에서 발견되며, 위치에 따라 뇌의 미세 아교 세포나 폐의 폐포대식세포와 같이 특정 이름으로 불리고 크기와 형태도 달라질 수 있다.[211] 대부분 조직에 상주하며 혈액 순환에서는 드물게 관찰되고, 조직 대식세포의 수명은 4일에서 15일 사이로 추정된다.[76][212]
대식세포의 주요 기능 중 하나인 식작용은 세균, 죽은 세포, 세포 잔해 등을 세포 안으로 삼키는 과정이다. 이 과정은 호중구에 비해 느리게 진행되어 몇 시간이 소요될 수 있으며, 대식세포는 많은 양의 물질을 탐식한다. 때로는 소화되지 않은 물질 일부를 조직으로 다시 방출하기도 하는데, 이는 다른 식세포를 불러 모으는 신호로 작용할 수 있다.[165][25] 대식세포 표면에는 옵소닌 수용체, 청소제 수용체, 톨유사수용체 등 다양한 수용체가 존재하여 외부 물질을 인식하고 결합한다.[142][12] 특히 옵소닌 수용체는 항체(면역글로불린 G)나 보체 단백질이 부착된 세균의 식작용을 효율적으로 만든다.[166][27] 톨유사수용체에 특정 분자(리간드)가 결합하면 식작용이 강화될 뿐만 아니라, 대식세포가 염증을 유발하는 사이토카인 분비를 촉진한다.[142][12]
평상시 조직 내에서 비교적 휴면 상태로 존재하는 대식세포는 주로 죽은 세포나 비감염성 물질을 제거하는 역할을 한다.[201][64] 그러나 감염이 발생하면 인터페론 감마(IFN-γ)와 같은 화학적 신호를 받아 활성화된다. 활성화된 대식세포는 MHC 클래스 II 분자 발현을 늘려 항원 제시 능력을 강화하고 병원체 살상 능력도 향상시킨다.[201][64] 침입한 병원체로부터 지질다당류 같은 직접적인 신호를 받으면 '과활성화' 상태가 되어 증식을 멈추고 병원체 제거에 집중하며, 크기와 식작용 속도가 증가한다.[201][64]
대식세포 활성화에는 림프구의 한 종류인 보조 T세포(Th 세포), 특히 Th1 세포가 중요한 역할을 한다. Th1 세포는 인터페론 감마를 분비하고 세포 표면에 CD40 리간드(CD40L) 단백질을 발현시켜 대식세포를 효과적으로 활성화한다.[213][77] 종양괴사인자 알파(TNF-α) 역시 중요한 활성화 신호이다.[211][75] 활성화된 대식세포는 일산화 질소(NO)와 활성 산소종을 생성하여 주변 미생물을 살상하고,[142][12] TNF-α, 인터루킨 1, 인터루킨 6 등의 사이토카인을 분비하여 염증 반응을 조절하고 다른 면역 세포들을 동원한다.[211][75] 이러한 활성화된 대식세포는 종양 세포 파괴에도 중요한 역할을 수행한다.[211][75] Th1 세포는 또한 다른 사이토카인을 분비하여 골수에서 단핵구와 호중구 생성을 촉진하거나 이들 세포가 감염 부위로 이동하도록 유도한다.[214][78]
대식세포는 수지상세포와 함께 전문적인 항원제시세포로 기능한다.[194][57] 식작용으로 외부 단백질(항원)을 섭취하면, 세포 내부에서 이를 펩타이드 조각으로 분해한다. 이 펩타이드 조각은 MHC 클래스 II 분자와 결합하여 세포 표면에 제시된다.[155][11] 이렇게 제시된 항원은 T 림프구(특히 보조 T세포)에 의해 인식되어 적응면역 반응을 개시하는 데 중요한 역할을 한다. 대식세포는 주로 감염 부위에 머무르며 항원을 제시하는 반면, 수지상세포는 림프절로 이동하여 더 광범위하게 면역 반응을 유도할 수 있다.[195][58]
7. 5. 호중구
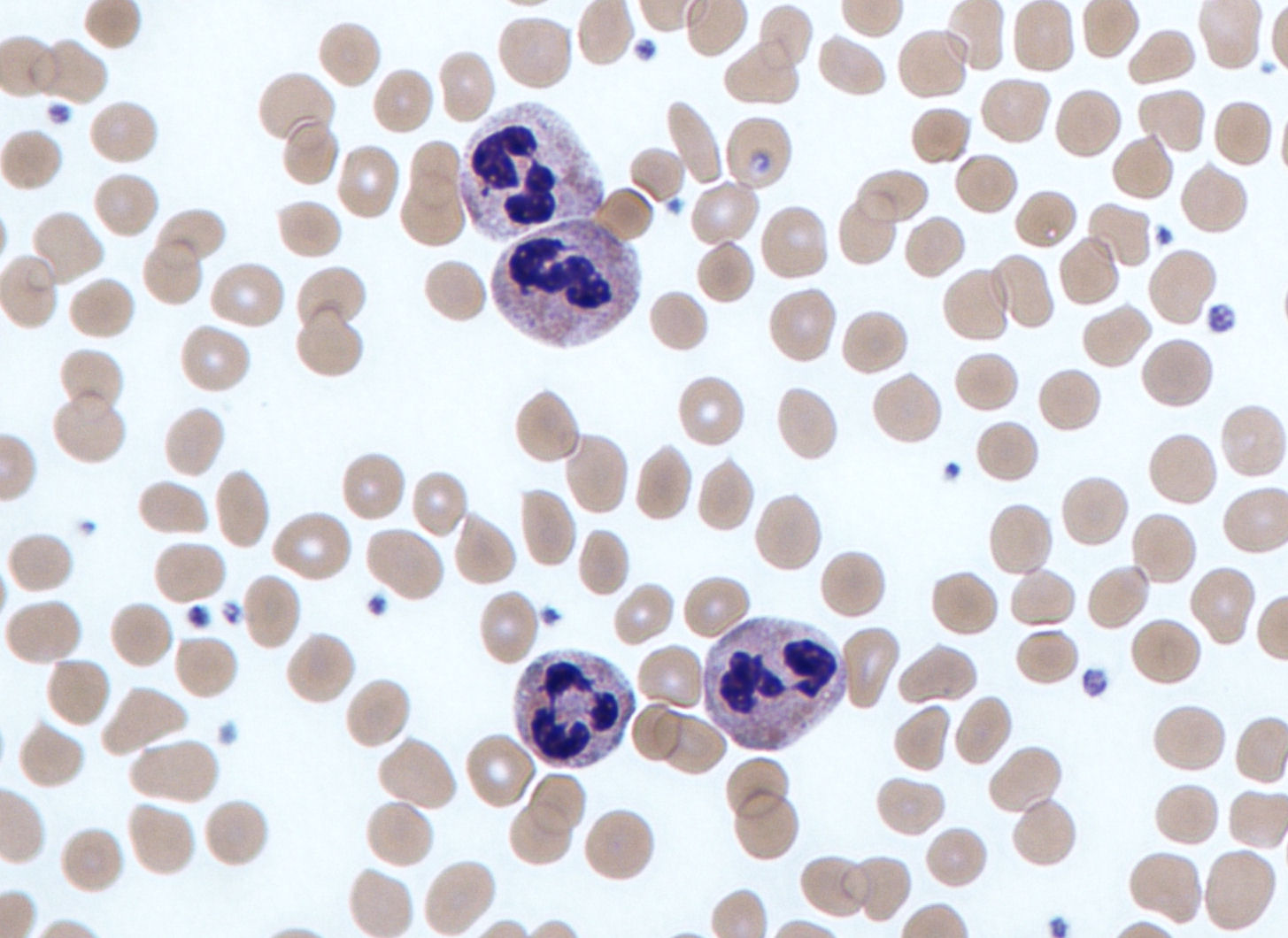
호중구는 혈류에서 가장 흔하게 발견되는 식세포 유형으로, 순환하는 전체 백혈구의 50~60%를 차지한다.[79][215] 사람 혈액 1리터에는 약 50억 개의 호중구가 있으며,[3][145] 직경은 약 10마이크로미터이다.[80][216] 수명은 약 5일 정도로 짧다.[38][176]
적절한 신호를 받으면 호중구는 혈액을 떠나 감염 부위에 도달하는 데 약 30분이 걸린다.[81][217] 감염이 발생하면 사이토카인과 같은 화학적 신호가 방출되어 호중구를 감염 부위로 유인한다.[12][142][153] 감염 신호는 혈관 내피세포가 셀렉틴 단백질을 생성하게 하여 호중구가 달라붙게 하고, 다른 신호는 내피세포 사이의 접합부를 느슨하게 만들어 혈관벽을 통과할 수 있게 한다. 주화성 과정을 통해 호중구는 사이토카인을 따라 감염 장소로 이동한다.[12][142]
호중구는 왕성한 식세포로서,[81] 항체와 보체 단백질로 둘러싸인 침입 물질, 손상된 세포, 세포 파편 등을 빠르게 삼킨다.[81][217] 사람 호중구가 세균을 식작용하는 데에는 평균 9분이 소요된다.[23][163] 식작용 후 호중구는 혈액으로 돌아가지 않고 고름 세포로 변하여 죽는다.[81][217] 감염 시 수백만 개의 호중구가 혈액에서 동원되지만 며칠 후 사멸한다.[69][205]
성숙한 호중구는 단핵구보다 작고 여러 조각으로 나뉜 세포핵을 가지고 있다. 각 조각은 염색질 필라멘트로 연결되어 있으며, 보통 2~5개의 핵 분절을 가진다.[82][218] 호중구는 일반적으로 성숙할 때까지 골수를 떠나지 않지만, 감염 중에는 전구 세포인 후골수구, 골수구, 전골수구가 골수에서 방출되기도 한다.[82][218]
호중구 내부의 과립은 단백질을 분해하고 세균을 죽이는 특성을 가지고 있다.[83][219] 호중구는 단핵구와 대식세포를 자극하는 물질을 분비할 수 있으며, 이 분비물은 식작용과 세포 내 살균에 관여하는 활성 산소종의 생성을 증가시킨다.[84][220] 호중구의 1차 과립에서 분비되는 물질은 면역글로불린 G(IgG) 항체로 코팅된 세균의 식작용을 자극한다.[85][221]
세균, 진균, 또는 활성화된 혈소판과 마주치면 호중구는 호중구 세포외 덫(NETs)이라는 그물 모양의 염색질 구조를 생성한다.[86][222] 주로 DNA로 구성된 NETs는 네토시스(netosis)라는 과정을 통해 병원체를 포획하여 사멸시킨다. NETs에 잡힌 병원체는 다양한 산화적 및 비산화적 메커니즘에 의해 죽게 된다.[86][222]
7. 6. 수지상세포
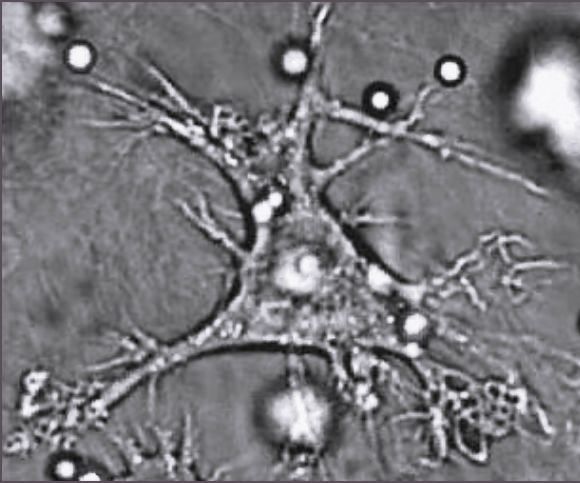
수지상세포는 나뭇가지처럼 길게 뻗은 돌기(수지상 돌기, dendrite영어)를 가진 특화된 항원제시세포이다.[87] 이 돌기는 미생물이나 다른 침입 물질을 식세포 작용하는 것을 돕는다.[88][89] 수지상세포는 주로 피부, 코 안쪽 점막, 폐, 위, 창자와 같이 외부 환경과 직접 접촉하는 조직에 존재한다.[90]
수지상세포는 항원제시에 중요한 역할을 한다. 외부 물질을 탐식하면 세포 내부에서 단백질(항원)을 작은 펩타이드 조각으로 분해한다. 이 펩타이드 조각은 주조직 적합성 복합체(MHC)라는 분자와 결합하여 세포 표면으로 이동하고, 면역계의 다른 세포, 특히 림프구에게 제시된다.[155] 수지상세포는 대식세포와 함께 전문적인 항원제시세포 중 하나이다.[194]
수지상세포가 활성화되면 성숙하여 림프절과 같은 림프 조직으로 이동한다.[91][58] 림프 조직에는 수많은 T세포와 B세포가 모여 있는데, 수지상세포는 이곳에서 T세포 및 B세포와 상호작용하며 적응면역 반응을 시작하고 조율하는 핵심적인 역할을 한다.[91] 성숙한 수지상세포는 보조 T세포와 세포독성 T세포를 효과적으로 활성화시킬 수 있다.[92] 활성화된 보조 T세포는 다시 대식세포나 B세포를 활성화시키는 데 기여한다. 또한, 수지상세포는 T세포가 특정 유형(세포독성 T세포 또는 보조 T세포)으로 분화하도록 유도함으로써 면역 반응의 방향을 결정하는 데 영향을 미친다.[88]
수지상세포는 면역 반응을 유도할 뿐만 아니라, 면역 관용을 촉진하여 우리 몸이 자신의 조직을 공격하는 자가 면역 반응을 억제하는 중요한 기능도 수행한다.[61] 만약 수지상세포가 우리 몸의 구성 성분(자기 항원)을 인식하게 되면, 이에 반응하는 림프구를 파괴하거나 비활성화시켜 면역 반응이 일어나지 않도록 한다. 이 과정을 면역 관용(immune tolerance)이라고 부른다.[60] 수지상세포는 흉선에서 자기 반응성 T세포를 제거하는 중추 관용과, 말초 조직에서 조절 T 세포 등을 통해 자기 반응성 T세포를 억제하는 말초 관용 모두에 기여한다.[62]
수지상세포는 골수의 조혈모세포에서 유래하는 전문 식세포 중 하나이며,[4] 혈액 속의 단핵구가 조직으로 이동하여 수지상세포로 분화하기도 한다.[71] 피부에 존재하는 랑게르한스 세포는 대표적인 수지상세포의 한 종류이다.
7. 7. 비만세포
비만세포는 톨유사수용체를 가지고 있으며 수지상세포, B세포, T세포와 상호작용하여 적응면역 기능을 중재하는 것을 돕는다.[229] 비만세포는 제2형 주조직 적합성 복합체 분자를 발현하여 항원제시에 참여할 수 있다. 그러나 항원제시에서 비만세포가 하는 정확한 기능은 아직 명확하게 밝혀지지 않았다.[230]비만세포는 그람 음성균(예: 살모넬라)을 탐식하고 살해할 수 있으며, 세균의 항원을 처리하는 능력도 가지고 있다.[231] 특히, 숙주 조직에 세균이 부착하는 데 중요한 역할을 하는 세균 표면의 섬모 단백질을 처리하는 데 특화되어 있다.[232][233] 이러한 기능 외에도, 비만세포는 염증 반응을 유도하는 사이토카인을 생산한다.[234] 사이토카인 분비는 미생물 파괴 과정에서 중요한 역할을 하는데, 이는 사이토카인이 감염 부위로 더 많은 식세포를 유인하여 면역 반응을 강화하기 때문이다.[231][235]
8. 비전문 식세포
죽어가는 세포와 외부로부터 유입된 유기체는 전문 식세포 이외의 다른 세포들에 의해서도 섭취될 수 있다.[237][101] 이러한 세포들에는 상피세포, 내피세포, 섬유아세포, 간엽 세포, 멜라닌 세포, 간세포 등이 있다.[106] 이들은 전문 식세포와 달리 식작용이 그들의 주된 기능이 아니라는 점을 강조하기 위해 비전문 식세포라고 불린다.[238][102] 예를 들어, 섬유아세포는 상처가 치유되거나 흉터가 재형성되는 과정에서 콜라겐을 식작용으로 처리하고, 외부 입자를 섭취하려는 시도를 하기도 한다.[239][103]
비전문 식세포는 전문 식세포에 비해 섭취할 수 있는 입자의 종류가 제한적이다. 이는 비전문 식세포의 표면에 효율적인 식세포 수용체, 특히 침입한 외부 물질이나 죽은 세포에 결합하는 옵소닌(항체와 보체 등)과 결합하는 수용체가 부족하기 때문이다.[146][4] 또한, 대부분의 비전문 식세포는 식작용에 대한 반응으로 활성 산소종(ROS)을 생산하지 않는다.[240][104]
9. 병원체의 식세포 회피와 저항
병원체는 숙주의 방어 체계를 뚫어야만 감염을 일으킬 수 있다. 특히 병원성을 가진 세균이나 원생동물(원충)은 식세포의 공격에 저항하기 위한 다양한 방법을 진화시켜 왔으며, 많은 병원체가 식세포에게 잡아먹힌 뒤에도 살아남아 오히려 그 안에서 복제하며 증식하기도 한다.[242][243][107][108] 이러한 생존 전략에는 식세포와의 접촉 자체를 피하거나, 식세포에 의한 탐식 과정을 방해하고, 식세포 내부에 침투하여 생존하거나, 심지어 식세포를 직접 공격하여 파괴하는 방법 등이 포함된다. 예를 들어 결핵균, 살모넬라균, 레지오넬라균 등 일부 세균은 식세포 내부의 소화 과정을 억제하며 생존하고,[242][107] 리슈만편모충과 같은 기생충은 식세포의 소화 작용에 저항하여 살아남기도 한다.[242][107]
9. 1. 접촉 회피
세균이 식세포와의 접촉을 피하는 방법은 여러 가지가 있다.[107][108]
첫째, 식세포가 이동할 수 없는 곳, 예를 들어 상처가 없는 피부 표면 같은 곳에서 자랄 수 있다. 둘째, 세균은 염증 반응을 억제할 수 있는데, 염증이 없으면 식세포가 감염에 제대로 반응하기 어렵다. 셋째, 일부 세균 종은 주화성 과정을 방해하여 식세포가 감염 장소로 이동하는 것을 막을 수 있다.[107] 넷째, 몇몇 세균은 자신이 숙주 세포의 일부(자기, self)인 것처럼 면역계를 속여 식세포를 회피하기도 한다. 매독의 원인균인 매독균(''Treponema pallidum'')은 자신의 표면을 피브로넥틴으로 덮어 식세포로부터 숨는다.[109][110] 피브로넥틴은 몸에서 자연적으로 만들어지며 상처 치유 과정에서 중요한 역할을 하는 물질이다.[110]
9. 2. 탐식 회피
병원체는 숙주 유기체의 방어 체계를 뚫고 지나가야만 성공적으로 감염을 일으킬 수 있다. 특히 병원성 세균과 원생동물은 식세포의 공격에 저항하기 위해 다양한 방법을 진화시켰으며, 많은 수가 식세포 내부에서 생존하고 증식하기도 한다.[107][108]세균이 식세포 작용을 회피하는 주요 방법 중 하나는 단백질이나 당으로 이루어진 협막(capsule)을 생산하여 세포 주변을 둘러싸는 것이다. 이 협막은 식작용 과정을 물리적으로 방해한다.[107][242] 예를 들어, 대장균(''Escherichia coli'') 표면의 K5 협막과 O75 O 항원,[246][111] 표피포도상구균(''Staphylococcus epidermidis'')의 세포밖다당류 협막이 이러한 역할을 한다.[247][112] 폐렴구균(''Streptococcus pneumoniae'')은 각기 다른 수준의 방어 능력을 제공하는 여러 종류의 협막을 생산할 수 있다.[248][113]
협막 외에도 특정 단백질을 생산하여 탐식을 회피하는 경우도 있다. A군 연쇄상구균은 M 단백질이나 선모 단백질 등을 생산하여 식세포에 의한 포획을 막는다.[248][113] 일부 단백질은 옵소닌화된 세균의 섭취를 방해하기도 한다. 예를 들어, 황색포도상구균(''Staphylococcus aureus'')은 단백질 A를 생산하여 항체가 세균 표면에 결합하는 것을 막음으로써 옵소닌의 효과를 감소시킨다.[249][114]
또한, 일부 세균은 독성 인자를 직접 분비하여 식세포의 기능을 방해한다. 예르시니아속(''Yersinia'')에 속하는 장병원성 세균들은 독성 인자인 YopH를 식세포 표면의 수용체에 결합시켜 식작용 능력을 저해한다.[250][115]
9. 3. 식세포 안에서 생존
병원체는 숙주의 방어 체계를 뚫어야만 감염에 성공할 수 있다. 병원성을 가진 세균과 원생동물(원충)은 식세포의 공격에 저항하는 다양한 방법을 발전시켜 왔으며, 많은 병원체가 식세포 안에서 살아남고 오히려 그 안에서 복제를 이어나갈 수 있다.[242][243][107][108]
세균은 면역계를 회피할 수 있는 공간인 식세포 안에서 생존하는 여러 방법을 개발했다.[251][116] 식세포 안으로 안전하게 들어가기 위해, 세균들은 인베이신(침입소)이라는 단백질을 발현한다.[251][116] 일단 식세포 안으로 들어가면, 세균은 세포질에 머무르며 파고리소좀 안에 있는 독성 화학 물질을 피하기도 한다.[252][117]
병원체가 식세포 내부에서 생존하는 주요 전략은 다음과 같다.
- 파고리소좀 형성 방해: 몇몇 세균은 파고솜과 리소좀이 융합하여 파고리소좀을 형성하는 과정을 막는다.[242][107] 예를 들어 결핵균( ''M. tuberculosis''), 살모넬라균, 레지오넬라균 등은 파고리소좀 형성을 억제하는 것으로 알려져 있다.
- 변형된 액포 형성: 리슈만편모충(''Leishmania'')과 같은 다른 병원체들은 식세포 안에서 크게 변형된 액포(공포)를 만드는데, 이는 병원체가 그 안에서 지속적으로 복제하며 생존하는 데 도움을 준다.[253][118]
- 파고리소좀 내부 생존: 일부 세균은 파고리소좀 안에서도 생존할 수 있다. 예를 들어, 황색포도상구균(''Staphylococcus aureus'')은 카탈레이스와 초과산화물 불균등화효소를 생산하여, 식세포가 세균을 죽이기 위해 사용하는 과산화 수소와 같은 화학 물질들을 분해한다.[254][119] 리슈만편모충속의 기생충 역시 파고리소좀 내에서 소화되는 것에 저항하여 생존할 수 있다.
- 파고솜 탈출: 세균은 파고리소좀이 형성되기 전에 파고솜에서 탈출하여 세포질로 이동할 수도 있다. 리스테리아 모노사이토제네스(''Listeria monocytogenes'')는 리스테리오라이신 O나 인지질분해효소 C(PLC)라는 효소를 이용하여 파고솜 벽에 구멍을 뚫고 빠져나간다.[255][120] 리케차 역시 파고솜을 통해 식세포 안으로 들어온 뒤 금방 파고솜을 탈출하여 식세포의 세포질 안에서 증식한다. 이때 파고솜 막을 뚫기 위해 인지질분해효소 A(PLA) 같은 효소가 사용되는 것으로 보인다.[242]
특정 세균은 여러 종류의 식세포를 감염 경로로 이용하기도 한다. 예를 들어, 결핵균(''M. tuberculosis'')은 호중구를 먼저 감염시킨 뒤, 이 감염된 호중구가 다시 대식세포에 의해 섭취되면서 대식세포를 감염시킨다.[121] 나병균(''M. leprae'')은 대식세포, 슈반 세포, 호중구 등을 감염시킬 수 있다.[121]
9. 4. 식세포 살해
세균은 식세포를 역으로 살해하는 다양한 방식들을 발전시켰다.[249][114] 예를 들어, 식세포의 세포막에 구멍을 뚫는 효소인 세포용해소를 사용하거나, 호중구의 과립이 파열되어 독성 물질이 방출되도록 만드는 스트렙토라이신과 류코시딘을 이용한다.[256][257][122][123] 또한, 식세포의 식작용에 필요한 ATP 공급을 감소시키는 외독소를 사용하기도 한다. 어떤 세균은 식세포에 섭취된 이후, 파고솜이나 파고리소좀의 막에 작용하는 독소를 방출하여 식세포의 다른 부분을 공격하고 결국 식세포를 죽게 만든다.[242][107]
9. 5. 세포 신호 방해
몇몇 세균은 사이토카인이나 다른 세포 신호를 방해하여 식세포가 침투한 병원체에 반응하는 것을 막는 생존 전략을 사용한다.[258][124] 예를 들어, 원충성 기생충인 톡소포자충(''Toxoplasma gondii''), 크루스파동편모충(''Trypanosoma cruzi''), 리슈만편모충(''Leishmania'') 등은 대식세포를 감염시키며, 각자 독특한 방법으로 대식세포의 기능을 방해한다.[258][124]
특히 리슈만편모충에 속하는 몇몇 종은 감염시킨 대식세포의 신호 전달 과정을 변경한다. 이를 통해 사이토카인이나 항미생물 분자(일산화 질소, 활성 산소 등)의 생산을 억제하고, 항원제시 과정을 막아 면역 반응을 회피한다.[259][125]
10. 식세포에 의한 숙주 손상
식세포, 특히 대식세포와 호중구는 감염을 조절하는 과정에서 단백질이나 작은 분자 형태의 염증 매개체를 방출하여 염증 반응의 중심적인 역할을 수행한다. 그러나 이러한 염증 과정은 숙주 조직에 손상을 입힐 수도 있다.[126] 일반적으로 식세포는 병원체를 삼켜 세포 내 소기관인 파고리소좀 안에서 독성 화학 물질을 이용해 파괴한다. 하지만 식세포가 목표 병원체를 삼키는 데 실패하면(이를 '좌절된 식작용'이라 부르기도 한다), 독성 물질이 세포 밖 환경으로 방출될 수 있다. 이 물질들은 숙주 세포에도 독성을 가지므로, 건강한 세포와 조직에 광범위한 손상을 일으킬 수 있다.[126]
호중구가 콩팥에서 과립 내용물을 방출하는 경우가 대표적인 예이다. 과립에 포함된 활성 산소종과 단백질분해효소는 숙주 세포 주변의 세포외기질을 분해하고 사구체 세포에 직접적인 손상을 입혀 혈액 여과 능력을 떨어뜨리고 세포 모양을 변화시킬 수 있다. 또한, 인지질분해효소의 생성물인 류코트리엔과 같은 물질은 이러한 손상을 더욱 악화시킨다. 류코트리엔 등은 더 많은 호중구를 감염 부위로 끌어들이는 주화성을 유발하며, 이 과정에서 사구체 세포는 이동하는 호중구의 부착 분자에 의해 추가적인 손상을 입을 수 있다. 결국 사구체 세포의 손상은 신부전으로 이어질 수 있다.[127][261]
호중구는 대부분의 급성 폐 손상 발생에도 핵심적인 역할을 한다.[128][262] 활성화된 호중구는 폐 환경으로 독성 과립 내용물을 방출하여 폐 조직을 손상시킨다.[129][263] 실험적으로 호중구 수를 줄이면 급성 폐 손상이 완화되는 것이 관찰되었지만,[130][264] 호중구 활동을 억제하는 치료법은 숙주를 감염에 매우 취약하게 만들기 때문에 임상적으로 적용하기 어렵다.[129][263] 또한, 세균이 만든 내독소의 방출, 패혈증, 외상, 알코올성 간염, 허혈, 급성 출혈로 인한 저혈량성 쇼크 등에 반응하여 호중구가 활성화되면 간 손상 및 기능 장애를 유발할 수도 있다.[131][265]
대식세포에서 방출되는 화학 물질 역시 숙주 조직에 손상을 줄 수 있다. 종양괴사인자 알파(TNF-α)는 대식세포가 분비하는 주요 물질 중 하나로, 평소에는 작은 혈관의 혈액 응고를 유도하여 감염이 퍼지는 것을 막는 역할을 한다.[132][266] 그러나 세균 감염이 혈액을 통해 전신으로 퍼지게 되면, TNF-α가 생명 유지에 필수적인 장기들로 방출되어 혈관 확장과 혈장 부피 감소를 유발할 수 있다. 이는 심각한 상태인 패혈성 쇼크로 이어질 수 있다. 패혈성 쇼크 상태에서는 TNF-α가 계속 작용하여 주요 장기로 가는 작은 혈관들을 막아 장기 기능 부전을 일으킬 수 있으며, 이는 사망에 이를 수도 있는 매우 위험한 상황이다.[67][153]
11. 진화적 기원
식작용은 진화 초기에[267][133] 단세포 진핵생물에서 처음 나타난 것으로 추정된다.[268][134] 식물이 갈라진 직후 후생동물 계통에서 분리된 단세포 원생생물인 아메바는 포유류의 식세포와 많은 기능을 공유한다.[268][134]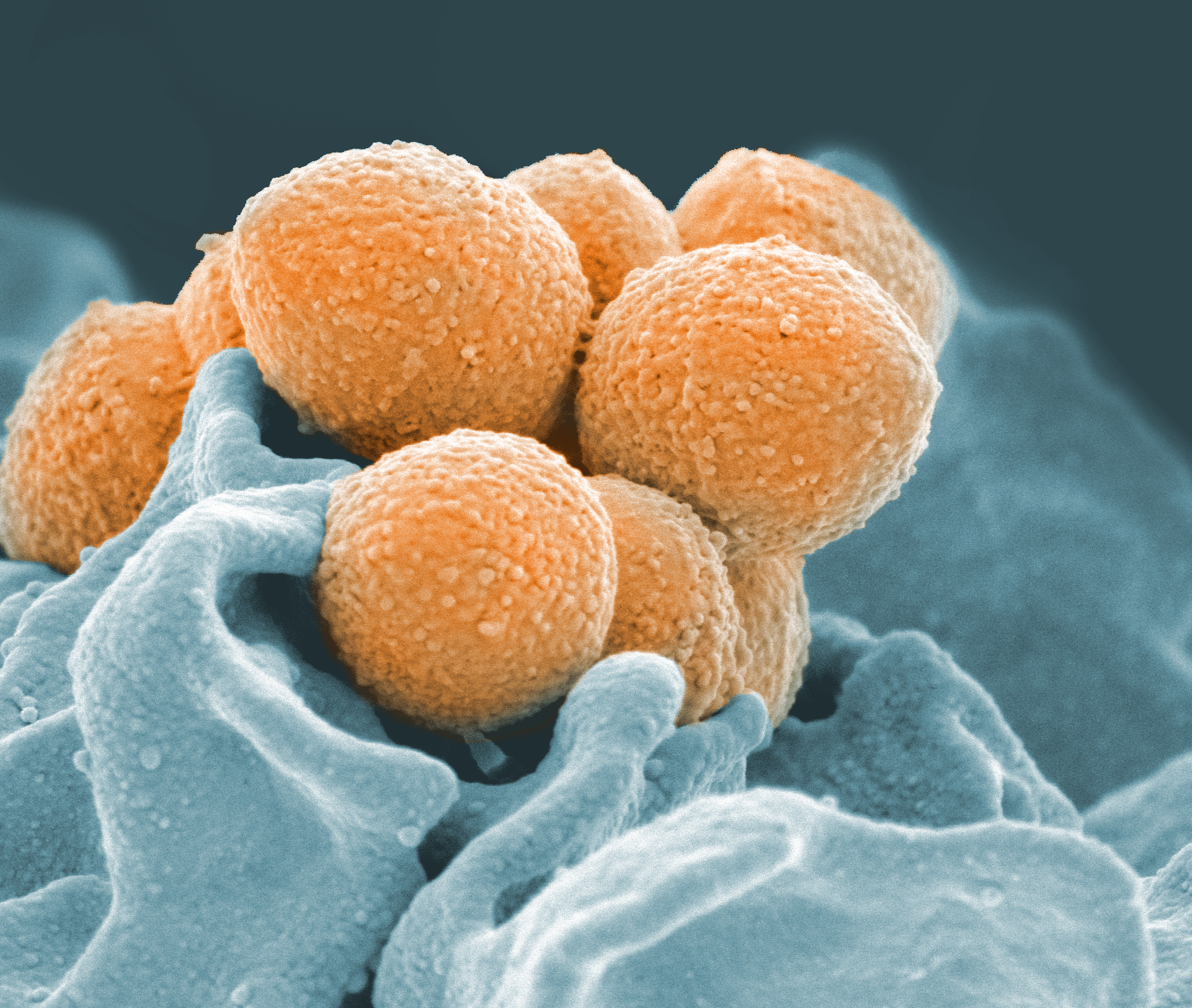
예를 들어, 딕티오스텔리움 디스코이데움(''Dictyostelium discoideum'')은 토양에 사는 아메바의 일종으로, 세균을 먹고 산다. 이 아메바는 동물의 식세포처럼 톨유사수용체를 통해 식작용으로 세균을 삼키며, 대식세포와 여러 생물학적 기능을 공유한다.[269][135] ''Dictyostelium discoideum''은 사회성을 보여주기도 하는데, 먹이가 부족하면 여러 세포가 모여 이동 가능한 다세포 형태인 슈도플라스모디움(pseudoplasmodium, slug) 또는 가소체(민달팽이 형태)를 형성한다. 이 다세포 유기체는 결국 환경 위험에 저항성이 있는 포자와 자실체를 만들어낸다. 자실체가 형성되기 전, 세포들은 슈도플라스모디움 형태로 이동하는데, 이 기간 동안 독소나 세균에 노출되면 포자 생산에 차질이 생겨 종의 생존을 위협할 수 있다. 이때 일부 아메바는 슈도플라스모디움 안을 돌아다니며 세균을 삼키고 독소를 흡수하는데, 이 아메바들은 결국 죽게 된다. 이들은 다른 아메바들과 유전적으로 동일하며, 다른 개체를 보호하기 위한 자기 희생은 고등 척추동물 면역계에서 식세포가 보이는 자기 희생과 유사하다. 이러한 사회성 아메바의 오래된 면역 기능은 세포성 섭식 메커니즘이 진화적으로 오래 보존되어 방어 기능으로 사용되고 있음을 시사한다.[270][136]
식세포는 해면동물부터 곤충류, 하등 및 고등 척추동물에 이르기까지 동물계 전반에 걸쳐 나타난다.[143][271][272][137][138] 아메바가 자기(self)와 비자기(non-self)를 구별하는 능력은 매우 중요하며, 많은 아메바 종의 면역계의 근본이 된다.[148][6]
참조
[1]
서적
[2]
서적
[3]
서적
[4]
웹사이트
Ilya Mechnikov
http://nobelprize.or[...]
2008-11-28
[5]
논문
Ilya Ilich Metchnikoff (1845–1915) and Paul Ehrlich (1854–1915): the centennial of the 1908 Nobel Prize in Physiology or Medicine
[6]
웹사이트
Evolution of the innate immune system.
https://www.ncbi.nlm[...]
2009-03-20
[7]
서적
[8]
서적
[9]
서적
[10]
논문
Antimicrobial reactive oxygen and nitrogen species: concepts and controversies
2004-10
[11]
서적
[12]
서적
[13]
논문
Immunology's Coming of Age
2019
[14]
서적
The Shorter Oxford English Dictionary
Oxford University Press (Guild Publishing)
[15]
논문
Medals, memoirs—and Metchnikoff
1998-04-01
[16]
웹사이트
Ilya Mechnikov
http://nobelprize.or[...]
The Nobel Foundation
2014-12-19
[17]
서적
[18]
서적
[19]
서적
[20]
서적
[21]
서적
[22]
논문
It takes a village: Phagocytes play a central role in fungal immunity
2019-05
[23]
논문
A single assay for measuring the rates of phagocytosis and bacterial killing by neutrophils
http://www.jleukbio.[...]
1994-02
[24]
서적
[25]
서적
[26]
서적
[27]
서적
[28]
논문
RNA Modifications Modulate Activation of Innate Toll-Like Receptors
2019-01
[29]
논문
The phagocytes: neutrophils and monocytes
2008-08
[30]
논문
Respiratory burst in human neutrophils
1999-12-17
[31]
논문
NADPH oxidase
[32]
논문
Myeloperoxidase
[33]
논문
Neutrophils, myeloperoxidase, and bronchiectasis in cystic fibrosis: green is not good
2004-09
[34]
서적
[35]
서적
[36]
간행물
Interferon-gamma: an overview of signals, mechanisms and functions
2004-02
[37]
서적
[38]
서적
[39]
간행물
Chronic granulomatous disease
2008-09
[40]
간행물
Chediak-Higashi syndrome
2008-01
[41]
서적
[42]
간행물
Laboratorial diagnosis of lymphocytic meningitis
2007-10
[43]
서적
[44]
간행물
Apoptosis in the pathogenesis and treatment of disease
[45]
서적
[46]
웹사이트
Apoptosis
http://www.merriam-w[...]
2014-12-19
[47]
간행물
Phosphatidylserine receptor is required for clearance of apoptotic cells
2003-11
[48]
간행물
Flippase and scramblase for phosphatidylserine exposure
2019-12
[49]
간행물
Cell corpse engulfment mediated by ''C. elegans'' phosphatidylserine receptor through CED-5 and CED-12
http://ntur.lib.ntu.[...]
[50]
간행물
Eat me or die
[51]
간행물
Phagosome maturation during the removal of apoptotic cells: receptors lead the way
2008-10
[52]
서적
[53]
서적
[54]
서적
[55]
서적
[56]
서적
[57]
웹사이트
Antigen Presenting Cells (APC)
http://pim.medicine.[...]
2014-12-19
[58]
서적
[59]
서적
[60]
서적
[61]
간행물
Dendritic cell-regulatory T-cell interactions control self-directed immunity
[62]
웹사이트
Dendritic Cells and Immune Tolerance
http://www.rockefell[...]
The Rockefeller University
2014-12-19
[63]
간행물
Immunological tolerance and autoimmunity
[64]
서적
[65]
서적
[66]
서적
[67]
문서
Janeway, Chapter: Induced innate responses to infection.
https://www.ncbi.nlm[...]
[68]
간행물
Leukocyte-epithelial interactions
2003-10
[69]
서적
[70]
서적
[71]
서적
[72]
서적
[73]
논문
Development and heterogeneity of macrophages and their related cells through their differentiation pathways
1996-07
[74]
논문
Cell size of alveolar macrophages: an interspecies comparison
1997-09
[75]
서적
[76]
서적
[77]
서적
[78]
서적
[79]
서적
Inflammation and Fever from Pathophysiology: Principles of Disease
Academic Electronic Press
2014-12-19
[80]
서적
[81]
서적
[82]
논문
Passive deformability of mature, immature, and active neutrophils in healthy and septicemic neonates
1998-12
[83]
서적
[84]
논문
Neutrophil secretion products regulate anti-bacterial activity in monocytes and macrophages
2008-01
[85]
논문
Neutrophil primary granule proteins HBP and HNP1-3 boost bacterial phagocytosis by human and murine macrophages
2008-10
[86]
논문
Neutrophil extracellular traps in immunity and disease
2018-02
[87]
논문
Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution
[88]
웹사이트
Dendritic Cells
https://web.archive.[...]
The Rockefeller University
2014-12-19
[89]
논문
Antigen presentation and T cell stimulation by dendritic cells
[90]
서적
[91]
논문
The instructive role of dendritic cells on T-cell responses
[92]
서적
[93]
논문
The immunoglobulin E-Toll-like receptor network
https://www.karger.c[...]
2014-12-19
[94]
논문
New developments in mast cell biology
2008-11
[95]
논문
Mast cell modulation of immune responses to bacteria
2001-02
[96]
논문
Type 1 fimbrial expression enhances ''Escherichia coli'' virulence for the urinary tract
1996-09
[97]
논문
Mast cells process bacterial Ags through a phagocytic route for class I MHC presentation to T cells
http://www.jimmunol.[...]
2014-12-19
[98]
논문
Mast cells in allergy and host defense
[99]
논문
The role of mast cells in the defence against pathogens
[100]
서적
[101]
논문
Innate apoptotic immunity: the calming touch of death
2008-07
[102]
논문
Phagocytic uptake of ''Encephalitozoon cuniculi'' by nonprofessional phagocytes
2000-12
[103]
논문
Involvement of actin filaments and integrins in the binding step in collagen phagocytosis by human fibroblasts
2001-01
[104]
논문
Professional and non-professional phagocytes: an introduction
1995-03
[105]
논문
Granulocytes: New Members of the Antigen-Presenting Cell Family
2017
[106]
논문
Cell-in-Cell Structures in the Liver: A Tale of Four E's
2020
[107]
웹사이트
Mechanisms of Bacterial Pathogenicity: Bacterial Defense Against Phagocytes
http://textbookofbac[...]
2008
2014-12-19
[108]
논문
Leishmania species: models of intracellular parasitism
http://jcs.biologist[...]
2014-12-19
[109]
논문
Bacterial avoidance of phagocytosis
2002-05
[110]
논문
Fibronectin fragmentation promotes alpha4beta1 integrin-mediated contraction of a fibrin-fibronectin provisional matrix
2005-09
[111]
논문
Loss of resistance to ingestion and phagocytic killing by O(-) and K(-) mutants of a uropathogenic ''Escherichia coli'' O75:K5 strain
1999-08
[112]
논문
A crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence
2004-12
[113]
논문
''Streptococcus pneumoniae'' capsular serotype 19F is more resistant to C3 deposition and less sensitive to opsonophagocytosis than serotype 6B
2009-02
[114]
논문
Immune evasion by staphylococci
2005-12
[115]
논문
Resistance to phagocytosis by Yersinia
2002-02
[116]
논문
Phagocytosis of bacterial pathogens: implications in the host response
2001-12
[117]
논문
A region of the ''Yersinia pseudotuberculosis'' invasin protein enhances integrin-mediated uptake into mammalian cells and promotes self-association
1999-03
[118]
논문
The biogenesis and properties of the parasitophorous vacuoles that harbour ''Leishmania'' in murine macrophages
1998-10
[119]
논문
Intracellular survival of ''Staphylococcus aureus'': correlating production of catalase and superoxide dismutase with levels of inflammatory cytokines
2008-07
[120]
논문
Cytolysin-dependent escape of the bacterium from the phagosome is required but not sufficient for induction of the Th1 immune response against Listeria monocytogenes infection: distinct role of Listeriolysin O determined by cytolysin gene replacement
2007-08
[121]
논문
Antimicrobial Activity of Neutrophils Against Mycobacteria
2021
[122]
논문
Mutational analysis of the group A streptococcal operon encoding streptolysin S and its virulence role in invasive infection
2005-05
[123]
논문
Staphylococcal cutaneous infections: invasion, evasion and aggression
2006-06
[124]
논문
Sabotage and exploitation in macrophages parasitized by intracellular protozoans
2005-01
[125]
논문
Subversion of host cell signalling by the protozoan parasite ''Leishmania''
2005
[126]
문서
Paoletti
[127]
논문
Neutrophils and renal failure
1999-08
[128]
논문
Neutrophil activation and acute lung injury
2001-02
[129]
논문
Neutrophil granule contents in the pathogenesis of lung injury
2006-01
[130]
논문
Neutrophils and acute lung injury
2003-04
[131]
논문
Host tissue damage by phagocytes
1997-12
[132]
논문
Porcine innate and adaptative immune responses to influenza and coronavirus infections
https://biblio.ugent[...]
2006-10
[133]
문서
[134]
논문
Eat, kill or die: when amoeba meets bacteria
2008-06
[135]
서적
International Review of Cell and Molecular Biology
2008
[136]
논문
Immune-like phagocyte activity in the social amoeba
2007-08
[137]
문서
[138]
논문
Development of macrophages of cyprinid fish
2009-04
[139]
웹사이트
食細胞
https://kotobank.jp/[...]
コトバンク
2013-03-01
[140]
웹사이트
マクロファージ
http://100.yahoo.co.[...]
Yahoo!百科事典
2013-03-01
[141]
서적
The Shorter Oxford English Dictionary
Oxford University Press (Guild Publishing)
[142]
간행물
[143]
간행물
[144]
간행물
[145]
간행물
[146]
웹사이트
Ilya Mechnikov
http://nobelprize.or[...]
2008-11-28
[147]
저널
Ilya Ilich Metchnikoff (1845–1915) and Paul Ehrlich (1854–1915): the centennial of the 1908 Nobel Prize in Physiology or Medicine
[148]
웹사이트
Evolution of the innate immune system.
https://www.ncbi.nlm[...]
Janeway
2009-03-20
[149]
간행물
[150]
간행물
[151]
간행물
[152]
저널
Apoptosis in the pathogenesis and treatment of disease
[153]
저널
Induced innate responses to infection
https://www.ncbi.nlm[...]
2001
[154]
저널
Antimicrobial reactive oxygen and nitrogen species: concepts and controversies
2004-10
[155]
간행물
[156]
저널
Immunology's Coming of Age
2019
[157]
저널
Medals, memoirs—and Metchnikoff
1998-04-01
[158]
웹인용
Ilya Mechnikov
http://nobelprize.or[...]
The Nobel Foundation
2014-12-19
[159]
간행물
[160]
간행물
[161]
간행물
[162]
간행물
[163]
저널
A single assay for measuring the rates of phagocytosis and bacterial killing by neutrophils
http://www.jleukbio.[...]
1994-02
[164]
간행물
[165]
간행물
[166]
간행물
[167]
저널
The phagocytes: neutrophils and monocytes
2008-08
[168]
저널
Respiratory burst in human neutrophils
1999-12-17
[169]
저널
NADPH oxidase
[170]
저널
Myeloperoxidase
[171]
저널
Neutrophils, myeloperoxidase, and bronchiectasis in cystic fibrosis: green is not good
2004-09
[172]
간행물
[173]
간행물
[174]
저널
Interferon-gamma: an overview of signals, mechanisms and functions
2004-02
[175]
간행물
[176]
서적
2019
[177]
논문
Chronic granulomatous disease
2008-09
[178]
논문
Chediak-Higashi syndrome
2008-01
[179]
서적
2019
[180]
논문
Laboratorial diagnosis of lymphocytic meningitis
2007-10
[181]
서적
2019
[182]
서적
2019
[183]
웹인용
Apoptosis
http://www.merriam-w[...]
2014-12-19
[184]
논문
Phosphatidylserine receptor is required for clearance of apoptotic cells
https://semanticscho[...]
2003-11
[185]
논문
Flippase and scramblase for phosphatidylserine exposure
2019-12
[186]
논문
Cell corpse engulfment mediated by ''C. elegans'' phosphatidylserine receptor through CED-5 and CED-12
https://semanticscho[...]
[187]
논문
Eat me or die
https://semanticscho[...]
[188]
논문
Phagosome maturation during the removal of apoptotic cells: receptors lead the way
2008-10
[189]
서적
2019
[190]
서적
2019
[191]
서적
2019
[192]
서적
2006
[193]
서적
2006
[194]
웹인용
Antigen Presenting Cells (APC)
http://pim.medicine.[...]
Dalhousie University
2014-12-19
[195]
서적
2006
[196]
서적
2019
[197]
서적
2006
[198]
논문
Dendritic cell-regulatory T-cell interactions control self-directed immunity
[199]
웹인용
Dendritic Cells and Immune Tolerance
http://www.rockefell[...]
The Rockefeller University
2014-12-19
[200]
논문
Immunological tolerance and autoimmunity
[201]
서적
2019
[202]
서적
2019
[203]
서적
2006
[204]
논문
Leukocyte-epithelial interactions
2003-10
[205]
서적
2019
[206]
서적
2005
[207]
서적
2006
[208]
서적
2019
[209]
논문
Development and heterogeneity of macrophages and their related cells through their differentiation pathways
1996-07
[210]
논문
Cell size of alveolar macrophages: an interspecies comparison
1997-09
[211]
간행물
[212]
간행물
[213]
간행물
[214]
간행물
[215]
서적
Inflammation and Fever from Pathophysiology: Principles of Disease
Academic Electronic Press
2014-12-19
[216]
간행물
[217]
간행물
[218]
저널
Passive deformability of mature, immature, and active neutrophils in healthy and septicemic neonates
1998-12
[219]
간행물
[220]
저널
Neutrophil secretion products regulate anti-bacterial activity in monocytes and macrophages
2008-01
[221]
저널
Neutrophil primary granule proteins HBP and HNP1-3 boost bacterial phagocytosis by human and murine macrophages
2008-10
[222]
저널
Neutrophil extracellular traps in immunity and disease
2018-02
[223]
저널
Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution
[224]
웹인용
Dendritic Cells
http://www.rockefell[...]
The Rockefeller University
2014-12-19
[225]
저널
Antigen presentation and T cell stimulation by dendritic cells
[226]
간행물
[227]
저널
The instructive role of dendritic cells on T-cell responses
[228]
간행물
[229]
저널
The immunoglobulin E-Toll-like receptor network
https://www.karger.c[...]
2014-12-19
[230]
저널
New developments in mast cell biology
2008-11
[231]
저널
Mast cell modulation of immune responses to bacteria
2001-02
[232]
저널
Type 1 fimbrial expression enhances ''Escherichia coli'' virulence for the urinary tract
1996-09
[233]
저널
Mast cells process bacterial Ags through a phagocytic route for class I MHC presentation to T cells
http://www.jimmunol.[...]
2014-12-19
[234]
저널
Mast cells in allergy and host defense
[235]
저널
The role of mast cells in the defence against pathogens
[236]
간행물
[237]
저널
Innate apoptotic immunity: the calming touch of death
2008-07
[238]
저널
Phagocytic uptake of ''Encephalitozoon cuniculi'' by nonprofessional phagocytes
2000-12
[239]
저널
Involvement of actin filaments and integrins in the binding step in collagen phagocytosis by human fibroblasts
2001-01
[240]
저널
Professional and non-professional phagocytes: an introduction
1995-03
[241]
저널
Granulocytes: New Members of the Antigen-Presenting Cell Family
2017
[242]
웹인용
Mechanisms of Bacterial Pathogenicity: Bacterial Defense Against Phagocytes
http://textbookofbac[...]
2008
2014-12-19
[243]
저널
Leishmania species: models of intracellular parasitism
http://jcs.biologist[...]
2014-12-19
[244]
저널
Bacterial avoidance of phagocytosis
2002-05
[245]
저널
Fibronectin fragmentation promotes alpha4beta1 integrin-mediated contraction of a fibrin-fibronectin provisional matrix
2005-09
[246]
논문
Loss of resistance to ingestion and phagocytic killing by O(-) and K(-) mutants of a uropathogenic ''Escherichia coli'' O75:K5 strain
1999-08
[247]
논문
A crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence
2004-12
[248]
논문
''Streptococcus pneumoniae'' capsular serotype 19F is more resistant to C3 deposition and less sensitive to opsonophagocytosis than serotype 6B
2009-02
[249]
논문
Immune evasion by staphylococci
2005-12
[250]
논문
Resistance to phagocytosis by Yersinia
2002-02
[251]
논문
Phagocytosis of bacterial pathogens: implications in the host response
2001-12
[252]
논문
A region of the ''Yersinia pseudotuberculosis'' invasin protein enhances integrin-mediated uptake into mammalian cells and promotes self-association
1999-03
[253]
논문
The biogenesis and properties of the parasitophorous vacuoles that harbour ''Leishmania'' in murine macrophages
1998-10
[254]
논문
Intracellular survival of ''Staphylococcus aureus'': correlating production of catalase and superoxide dismutase with levels of inflammatory cytokines
2008-07
[255]
논문
Cytolysin-dependent escape of the bacterium from the phagosome is required but not sufficient for induction of the Th1 immune response against Listeria monocytogenes infection: distinct role of Listeriolysin O determined by cytolysin gene replacement
2007-08
[256]
논문
Mutational analysis of the group A streptococcal operon encoding streptolysin S and its virulence role in invasive infection
2005-05
[257]
논문
Staphylococcal cutaneous infections: invasion, evasion and aggression
2006-06
[258]
논문
Sabotage and exploitation in macrophages parasitized by intracellular protozoans
2005-01
[259]
논문
Subversion of host cell signalling by the protozoan parasite ''Leishmania''
2005
[260]
간행물
[261]
논문
Neutrophils and renal failure
1999-08
[262]
논문
Neutrophil activation and acute lung injury
2001-02
[263]
논문
Neutrophil granule contents in the pathogenesis of lung injury
2006-01
[264]
논문
Neutrophils and acute lung injury
https://semanticscho[...]
2003-04
[265]
논문
Host tissue damage by phagocytes
1997-12
[266]
논문
Porcine innate and adaptative immune responses to influenza and coronavirus infections
https://biblio.ugent[...]
2006-10
[267]
간행물
[268]
논문
Eat, kill or die: when amoeba meets bacteria
2008-06
[269]
서적
Phagocytosis and host-pathogen interactions in Dictyostelium with a look at macrophages
2008
[270]
논문
Immune-like phagocyte activity in the social amoeba
2007-08
[271]
간행물
[272]
논문
Development of macrophages of cyprinid fish
2009-04
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com